В данном материале подробно разбирается строение пародонта с точки зрения клинической анатомии и гистологии.
Общее строение пародонта
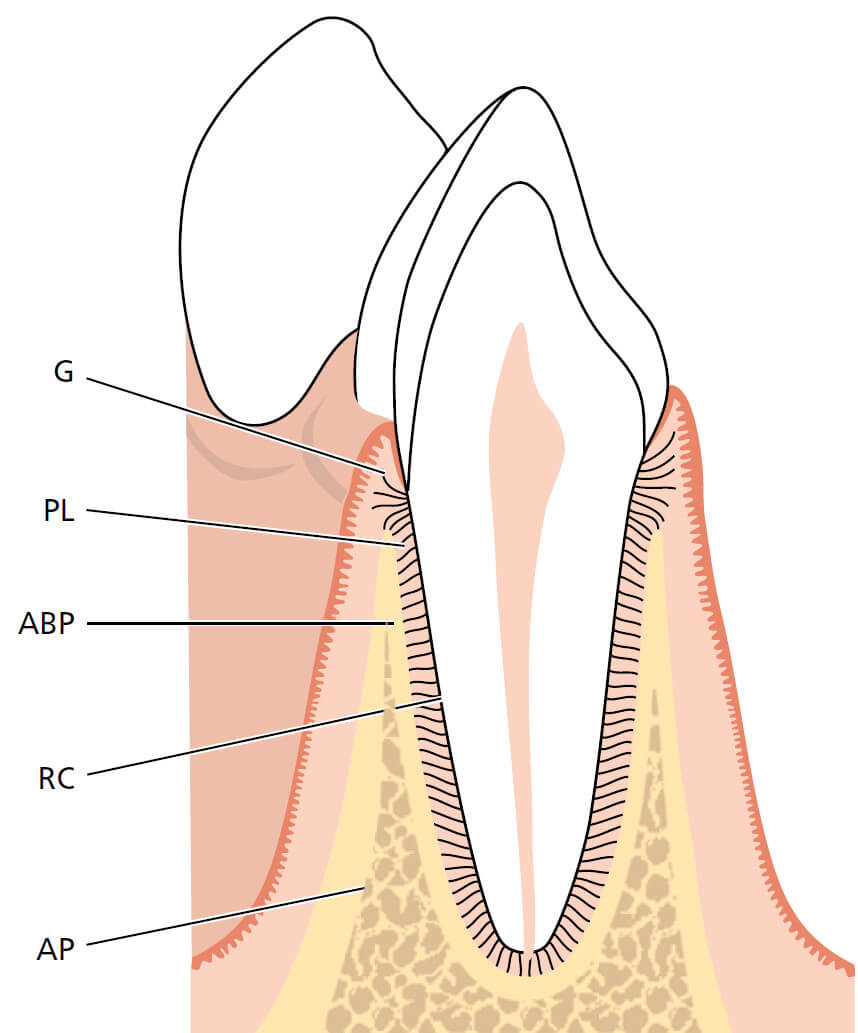
Пародонт (paro (perio) = вокруг, odontos = зуб) объединяет следующие ткани: (1) десну (G), (2) периодонтальную связку (PL), (3) корневой цемент (RC) и (4) собственно альвеолярную кость (АBP) (рис. 1).
АBP выстилает альвеолу зуба и неразрывно связана с альвеолярной костью; на рентгенограмме она может выглядеть как пластинка твердой мозговой оболочки. Альвеолярный отросток, который простирается от базальной кости верхней и нижней челюсти состоит из альвеолярной кости и собственно альвеолярной кости.
Основная функция пародонта заключается в прикреплении зуба к костной ткани челюстей и поддержании целостности поверхности жевательной слизистой оболочки полости рта.
Пародонт, также называемый “опорным аппаратом” или “опорными тканями зубов”, представляет собой развивающуюся, биологическую и функциональную единицу, которая с возрастом претерпевает определенные изменения и, кроме того, подвергается морфологическим изменениям, связанным с функциональными изменениями и изменениями в ротовой среде.
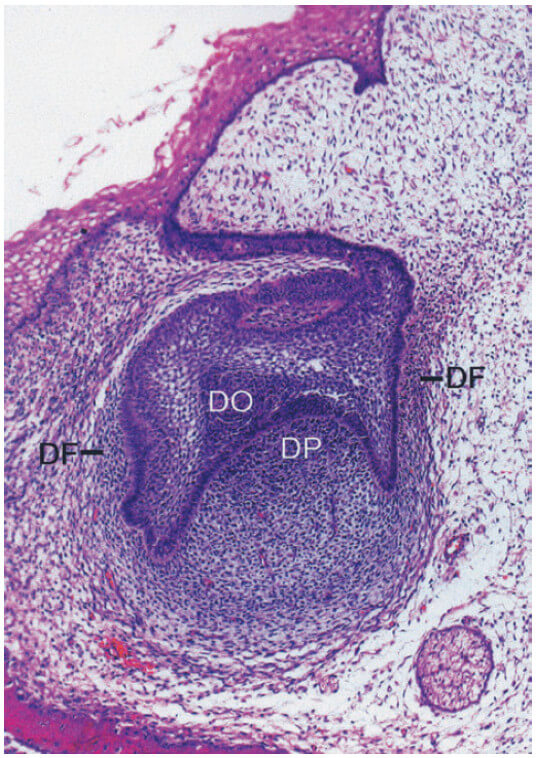
Развитие тканей пародонта происходит во время развития и формирования зубов. Этот процесс начинается на ранней стадии эмбриональной фазы, когда клетки из нервного гребня (из нервной трубки эмбриона) мигрируют в первичную ветвящуюся дугу. В этом положении клетки нервного гребня образуют полосу эктомезенхимы под эпителием стоматодея (примитивной полости рта). После того, как незафиксированные клетки нервного гребня достигли своего местоположения в челюстном пространстве, эпителий стоматодеума высвобождает факторы, которые инициируют эпителиально-эктомезенхимальные взаимодействия. После того, как эти взаимодействия произошли, эктомезенхима принимает доминирующую роль в дальнейшем развитии. После образования зубной пластинки начинается ряд процессов (стадия бутона, стадия колпачка, стадия колокола с развитием корня), которые приводят к образованию зуба и окружающих его тканей пародонта, включая собственно альвеолярную кость. Во время стадии колпачка происходит конденсация эктомезенхимальных клеток по отношению к зубному эпителию (рис. 2) (зубной орган [DO]), образуя зубной сосочек (DP), который дает начало дентину и пульпе, и зубной фолликул (DF), который дает начало опорным тканям пародонта (рис. ниже). Решающая роль, которую играет эктомезенхима в этом процессе, дополнительно устанавливается тем фактом, что ткань зубного сосочка, по-видимому, также определяет форму и вид зуба.
Если зачаток зуба в стадии развития колокола рассекается и пересаживается в эктопическое место (например, соединительной ткани или передней камеры глаза), процесс формирования зуба продолжается. Образуются коронка и корень, а также опорные структуры (цемент, периодонтальная связка и тонкая пластинка собственно альвеолярной кости). Такие эксперименты подвтерждают, что вся информация, необходимая для формирования зуба и его прикрепляющего аппарата находится в тканях зубного органа и окружающей эктомезенхиме. Зубной орган является формообразующим органом эмали, зубной сосочек–формообразующим органом комплекса «дентин-пульпа», а зубной фолликул-формообразующим органом прикрепительного аппарата (цемент, периодонтальная связка и собственно альвеолярная кость).
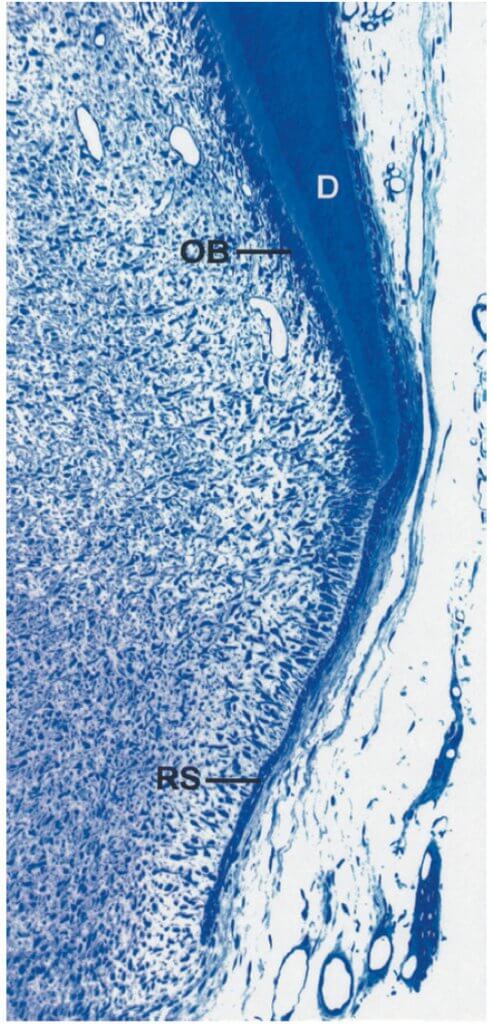
Развитие корня и поддерживающих тканей пародонта следует за развитием коронки. Эпителиальные клетки наружного и внутреннего зубного эпителия (зубного органа) пролиферируют в апикальном направлении, образуя двойной слой клеток, называемый эпителиальной оболочкой корня Гертвига (RS). Одонтобласты (ОBs) образующие дентин корня дифференцируются от эктомезенхимальных клеток в зубном сосочке под индуктивным воздействием внутренних эпителиоцитов (рис. ниже). Дентин (D) продолжает формироваться в апикальном направлении, производящее каркас корня. В процессе формирования корня развиваются опорные ткани пародонта, в том числе бесклеточный цемент. Некоторые данные в цементогенезе до сих пор остаются неясными, но постепенно появляется следующая концепция.
В начале формирования дентина, внутренние клетки эпителиальной оболочки корня Гертвига синтезируют и секретируют связанные с эмалью белки, вероятно, принадлежащие к семейству амелогенинов. В конце всего этого периода, эпителиальная оболочка корня становится фенестрированной, и эктомезенхимальные клетки из зубного фолликула проникают через эти фенестрации и контактируют с поверхностью корня. Эктомезенхимальные клетки, контактирующие с белками, связанными с эмалью, дифференцируются в цементобласты и начинают образовывать цементоид. Этот цементоид представляет собой органическую матрицу цемента и состоит из измельченного вещества и коллагеновых волокон, которые смешиваются с коллагеновыми волокнами в еще не полностью минерализованном наружном слое дентина. Предполагается, что цемент становится прочно прикрепленным к дентину через эти взаимодействия волокон.
Образование клеточного цемента, который часто покрывает апикальную треть зубных корней, отличается от образования бесклеточного цемента тем, что некоторые из цементных бластов внедряются в цемент.
Остальные части пародонта образованы эктомезенхимальными клетками от зубного фолликула латерально к цементу. Некоторые из них дифференцируются в периодонтальные фибробласты и образуют волокна периодонтальной связки, в то время как другие становятся остеобластами и формируют собственно альвеолярную кость, в которой закреплены волокна периодонта. Другими словами, первичная альвеолярная стенка также является эктомезенхимальным продуктом. Вполне вероятно, но до сих пор неопровержимо доказано, что эктомезенхимальные клетки остаются в зрелом периодонте и принимают участие в жизнедеятельности этой ткани.
Анатомия десны. Макроскопическая картина
Слизистая оболочка полости рта (слизистая оболочка) непрерывна связана с кожей губ и слизистой оболочкой мягкого неба и глотки. Слизистая оболочка полости рта состоит из (1) жевательной оболочки, которая включает десну и слизистую твердого неба; (2) специализированной слизистой оболочки, которая покрывает спинку языка; и (3) оставшейся части слизистой оболочкой.
Десна — это та часть жевательной слизистой оболочки, которая покрывает альвеолярный отросток и окружает пришеечную часть зубов (рис. 4). Она состоит из эпителиального слоя и нижележащего слоя соединительной ткани называемый собственная пластина. Десна приобретает свою окончательную форму и текстуру вместе с прорезыванием зубов.
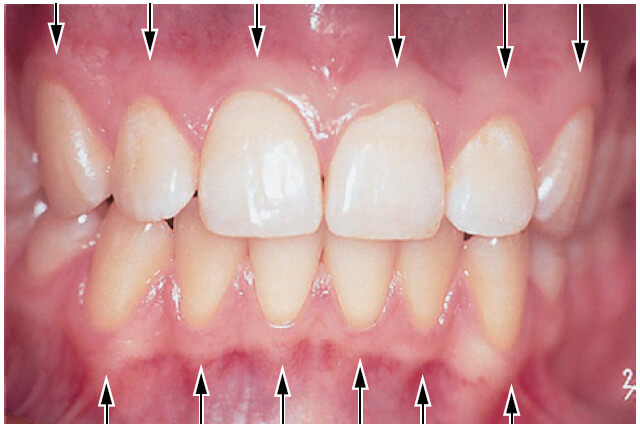
В направлении коронки зуба десна имеет кораллово-розовый цвет и заканчивается свободным десневым краем, который имеет зубчатый контур. В апикальном направлении десна непрерывна связана с рыхлой, темно-красной альвеолярной слизистой оболочкой (выстилающей слизистую оболочку), от которой десна отделена обычно легко узнаваемой границей, называемой либо мукогингивальным соединением (стрелки), либо мукогингивальной линией.
На твердом нёбе нет мукогингивальной линии, так как оно и верхнечелюстной альвеолярный отросток покрыты одним и тем же типом жевательной слизистой оболочки (рис. 5).
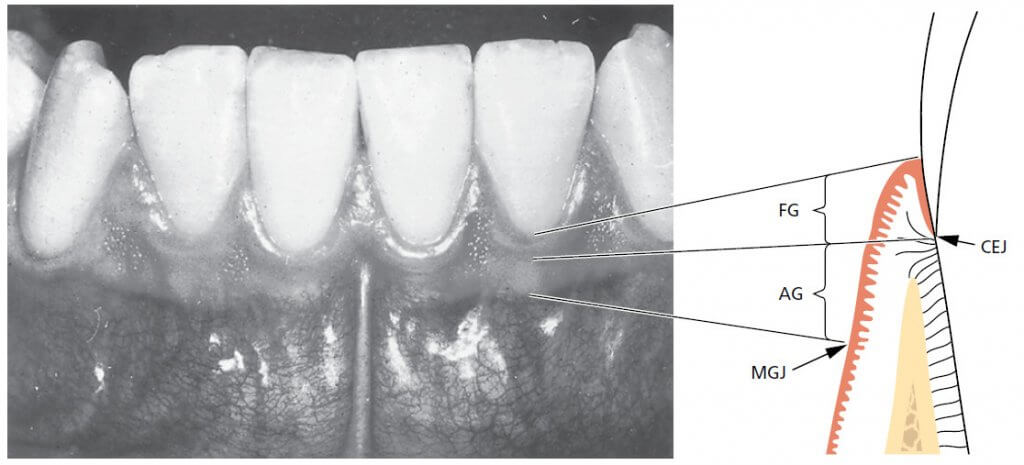
Три части десны могут быть определены следующим образом (рис. 6):
- Свободная десна (FG)
- Межзубная десна
- Прикрепленная десна (АG).
Свободная десна кораллово-розового цвета, имеет тусклую поверхность и твердую консистенцию. Она включает в себя десневую ткань на вестибулярной и язычно-небной сторонах зубов. На вестибулярной и язычной поверхностях свободная десна простирается от десневого края в апикальном направлении до свободной десневой борозды, которая расположена на уровне, соответствующем уровню цементоэмалевого соединения (CEJ). Прикрепленная десна отделена за счет мукогингивальным соединением (MGJ) в апикальном направлении.
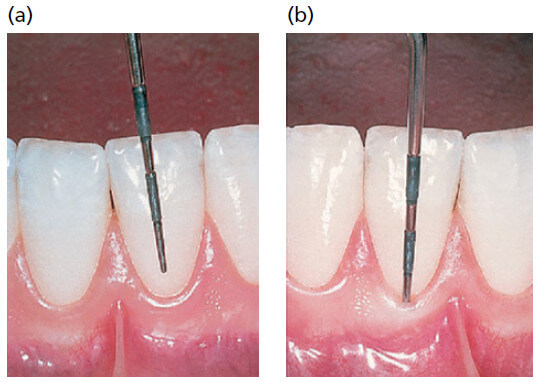
Свободный край десны часто закругляется таким образом, что между зубом и десной образуется небольшая инвагинация или борозда.
Когда в эту инвагинацию вводится пародонтальный зонд и продвигается далее апикально (рис. 7), по направлению к CEJ, десневая ткань отделяется от зуба и искусственно открывается “десневой карман” или “десневая щель”. Таким образом, в нормальной или клинически здоровой десне фактически отсутствует “десневой карман” или “десневая щель”, но десна находится в тесном контакте с поверхностью эмали. На части b показано, как пародонтальный зонд был введен в промежуток зуб–десна и “десневая щель” искусственно открыта приблизительно до уровня CEJ. После полного прорезывания зуба свободный край десны располагается на поверхности эмали примерно на 1,5 — 2 мм коронкой к CEJ.
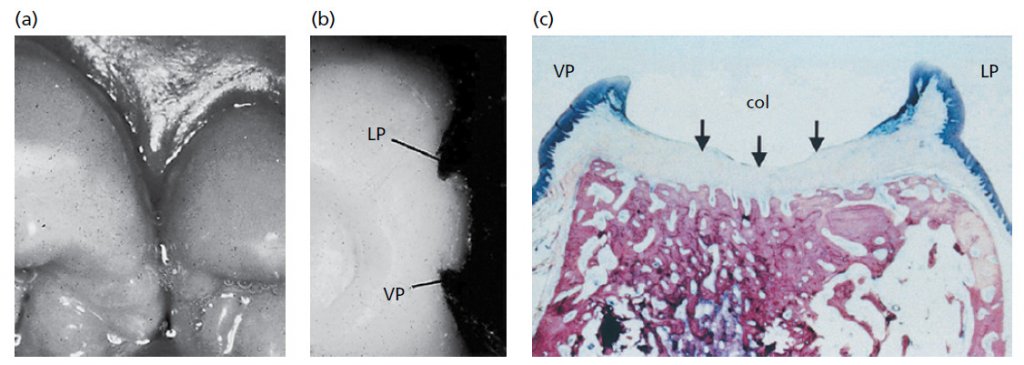
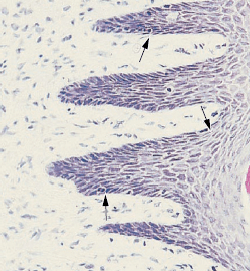
Форма межзубной десны (межзубный сосочек) определяется контактными отношениями между зубов, шириной прилегающих поверхностей зубов и ходом CEJ (рис. 8). В передних отделах зубного ряда межзубной сосочек имеет пирамидальную форму (b), в то время как в молярных отделах сосочки более плоские в букко-лингвальном направлении (a). Из-за наличия межзубных сосочков свободный десневой край более или менее выражен по отношению к зубному ряду.
В премолярных/молярных областях зубного ряда (рис. 9), зубы имеют аппроксимальные контактные поверхности (a), а не точки соприкосновения. Поскольку форма межзубного сосочка соответствует контуру межзубных контактных поверхностей, то в премолярной и молярной областях, как показано на рис.б, где удален дистальный зуб, образуется вогнутость – «седло». Таким образом, межзубные сосочки в этих областях часто имеют одну вестибулярную (VP) и одну язычную/небную часть (LP), разделенные областью седла. Область его, как показано на гистологическом срезе (с), покрыта тонким некератинизированным эпителием (стрелки). Этот эпителий имеет много общих черт с соединительным эпителием.
Прикрепленная десна является разграниченной во фронтальном направлении свободной десневой борозды (GG) или, если такой борозды нет, то в горизонтальном направлении на уровне CEJ. При клинических обследованиях было отмечено, что свободная десневая борозда присутствует только у 30-40% взрослых.
Свободная десневая борозда часто наиболее выражена на вестибулярной стороне зубов, чаще всего встречаясь в резцовой и премолярной областях зубов нижней челюсти, и реже всего в нижнечелюстных молярах и верхнечелюстных премолярах.
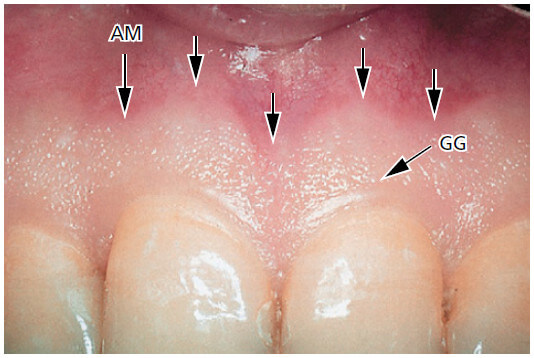
Прикрепленная десна простирается в апикальном направлении до мукогингивального соединения (стрелки), где она соединяется непрерывно с альвеолярной (выстилающей) слизистой оболочкой (АМ). Она имеет твердую текстуру, кораллово-розовый цвет, и часто имеет небольшие углубления на поверхности. Углубления, называемые “зернистостью”, дают вид апельсиновой корки.
Десна прочно прикреплена к нижележащей альвеолярной кости и цементу соединительнотканными волокнами и поэтому сравнительно неподвижна по отношению к нижележащей ткани. Более темная красная слизистая оболочка альвеол (АМ) расположенная апикально к мукогингивальному соединению, с другой стороны, слабо связана с нижележащей костью. Поэтому, в отличие от прикрепленной десны, альвеолярная слизистая оболочка подвижна по отношению к подлежащей ткани.
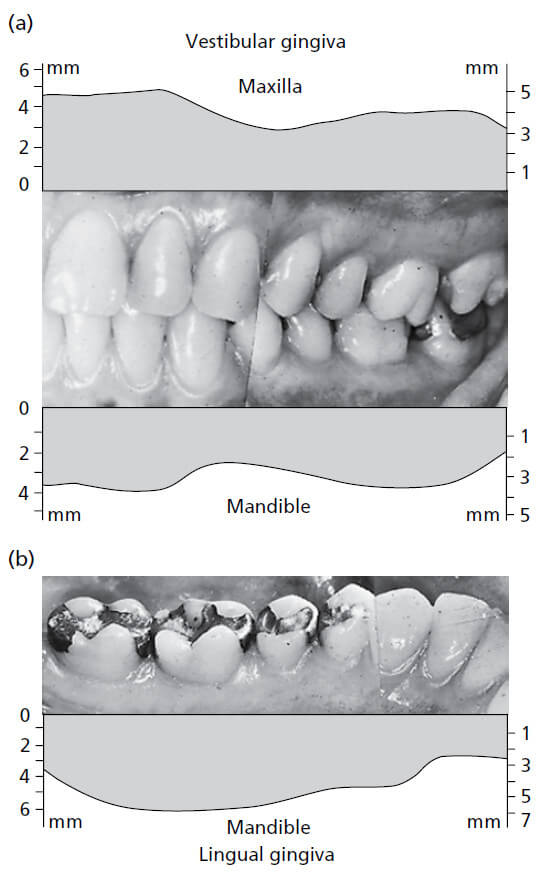
На рис. 11 показано, как изменяется ширина десны в разных частях зубного ряда. На верхней челюсти (а), вестибулярная десна обычно самая широкая в области резцов и самая узкая рядом с премолярами. На нижней челюсти (b), десна на язычной стороне особенно узка в области резцов и широка в молярной области. Диапазон ширины составляет 1–9мм.
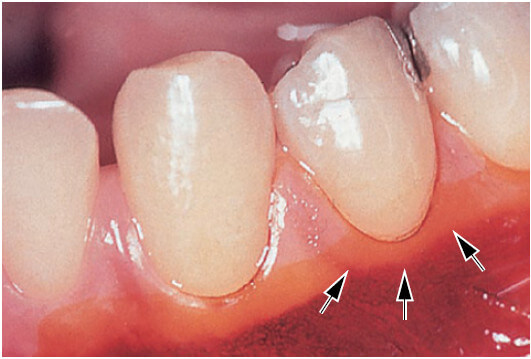
На рис. 12 показана область в премолярной области нижней челюсти, где десна чрезвычайно узкая. Стрелки указывают на расположение слизистой оболочки десны. Слизистая оболочка была окрашена раствором йода, чтобы более точно отличить десну от альвеолярной слизистой оболочки.
На рис. 13 представлен результат исследования, в котором оценивалась ширина прикрепленной десны и соотносилась с возрастом обследованных пациентов. Оказалось, что десна у 40-50‐летних значительно шире, чем у 20-30 — летних. Это наблюдение указывает на то, что ширина десны имеет тенденцию увеличиваться с возрастом. Поскольку слизисто-десневое соединение остается стабильным на протяжении всей жизни по отношению к нижней границе нижней челюсти, увеличение ширины десны может свидетельствовать о том, что зубы в результате окклюзионного износа медленно прорезываются на протяжении всей жизни.
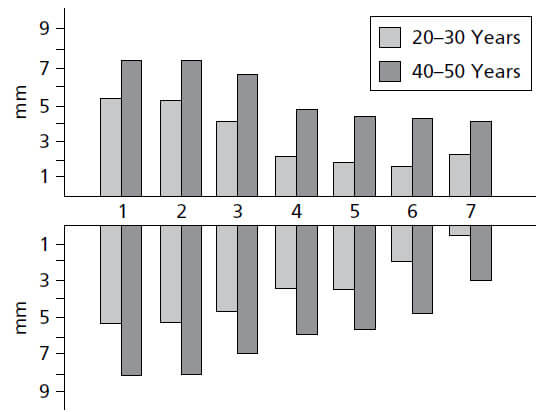
Анатомия десны. Микроскопическая картина: эпителий
Схематический рисунок гистологического среза (b), описывающий строение десны и область контакта между десной и эмалью (Е).
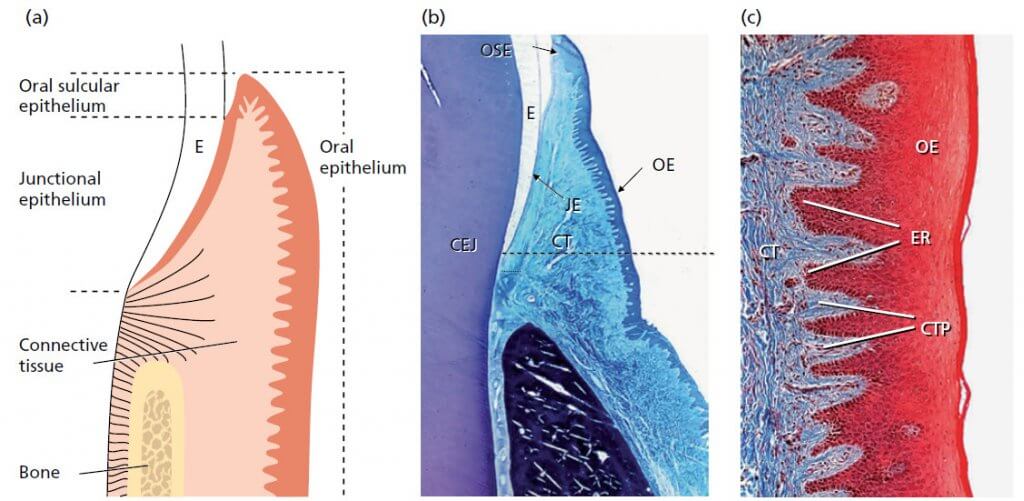
b — свободная десна включает в себя все эпителиальные и соединительнотканные структуры (СТ), расположенные коронарно к горизонтальной линии, расположенной на уровне цементо-эмалевого соединения (CEJ). Эпителий, покрывающий свободную десну, может быть дифференцирован следующим образом:
- Эпителий ротовой полости (ОE), обращенный к полости рта.
- Эпителий борозды (ОSE), который обращен к зубу, не соприкасается с поверхностью зуба.
- Соединительный эпителий (JE), который обеспечивает контакт между десной и зубом.
с — граница между эпителием полости рта (ОЕ) и подлежащей соединительной тканью (СТ) имеет волнообразный ход. Участки соединительной ткани, выступающие в эпителий, называются соединительнотканными сосочками (CTP) и отделены друг от друга эпителиальными гребнями – так называемыми ретикулярными колышками (ER). В нормальной, невоспаленной десне на границе между соединительным эпителием и подлежащей ему соединительной тканью отсутствуют ретикулярные колышки и соединительнотканные сосочки (b). Таким образом, характерной морфологической особенностью эпителия ротовой полости и бороздчатого эпителия является наличие ретикулярных колышков: эти структуры отсутствуют в соединительном эпителии.
На рис. 15 представлена модель, построенная на основе увеличенных гистологических срезов, показывающая подповерхностный слой ротового эпителия десны после удаления соединительной ткани. На поверхности эпителия полости рта (то есть на поверхности эпителия, обращенной к соединительной ткани) имеется несколько углублений, соответствующих соединительнотканным сосочкам (см. рис. ниже), которые выступают в эпителий. Можно видеть, что эпителиальные выступы, которые в гистологических срезах отделяют сосочки соединительной ткани, образуют непрерывную систему эпителиальных гребней.

На рис. 16 представлена модель соединительной ткани, соответствующая модели эпителия, показанной на рис. ранее. Эпителий был удален, тем самым сделав вестибулярную сторону десневой соединительной ткани видимым. Обратите внимание на соединительнотканные сосочки, выступающие в пространство, которое было заполнено эпителием рта (ОE) на рис. выше и эпителием борозды (ОSE) на задней стороне модели.

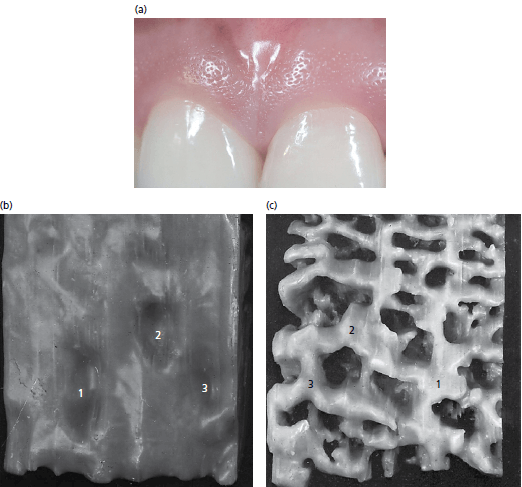
На рисунке 17:
а — у большинства взрослых прикрепленная десна имеет зернистость на поверхности. Фотография показывает случай, когда это зернистость бросается в глаза.
b — представлена увеличенная модель наружной поверхности ротового эпителия прикрепленной десны. Поверхность имеет мельчайшие углубления (1-3) которые придают десне характерный зернистый вид.
с — показана фотография подповерхностного слоя (то есть поверхности эпителия, обращенной к соединительной ткани) модели, показанной на рис. b. Подповерхностный слой эпителия характеризуется наличием эпителиальных гребней, которые сливаются на различных уровнях (1-3). Углубления видны на наружной поверхности эпителия (1-3) и соответствуют этим местам слияния (1-3) между эпителиальными гребнями. Таким образом, углубления на поверхности десны возникают в местах слияния между различными эпителиальными гребнями.
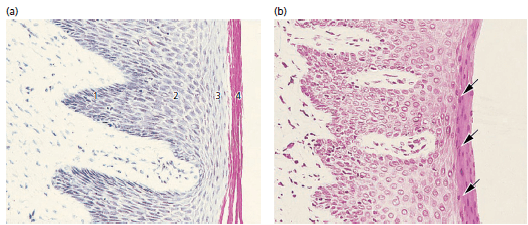
На рисунке 18:
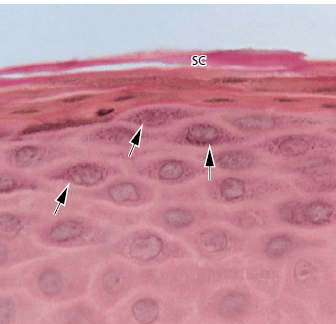
а — часть эпителия полости рта, покрывающая свободную десну, проиллюстрирована на этой микрофотографии. Эпителий ротовой полости представляет собой ороговевший, слоистый, плоский эпителий, который в зависимости от степени дифференцировки кератино продуцирующих клеток можно разделить на следующие клеточные слои:
- Базальный слой (stratum basale или stratum germinativum)
- Шиповатый клеточный слой (stratum spinosum)
- Зернистый клеточный слой (stratum granulosum)
- Ороговевший клеточный слой (stratum corneum)
Следует отметить, что в этом разделе ядра клеток отсутствуют во внешних клеточных слоях. Такой эпителий называется ортокератинизированным. Однако часто, клетки рогового слоя эпителия десны человека содержат остатки ядер, как показано на рис. b (стрелки). В таком случае эпителий обозначается как паракератинизированный.
Помимо клеток, продуцирующих кератин, которые составляют около 90% от общей популяции клеток, эпителий полости рта содержит следующие типы клеток:
- Меланоциты
- Клетки Лангерганса
- Клетки Меркеля
- Воспалительные клетки
Эти типы клеток часто звездчатые и имеют цитоплазматические расширения различного размера и внешнего вида. Их также называют “прозрачными клетками”, так как в гистологических срезах зона вокруг их ядер кажется светлее, чем в окружающих кератино продуцирующих клетках.
На микрофотографии (рис. 19) показаны “прозрачные клетки” (стрелки) расположенные в базальном слое эпителия рта или вблизи него. За исключением клеток Меркеля, эти “прозрачные клетки”, которые не производят кератин, не имеют десмосомной связи с соседними клетками. Меланоциты являются пигмент синтезирующими клетками и отвечают за пигментацию и отложения меланина, которая иногда наблюдается на десне. Однако как у светлых, так и у смуглых людей в эпителии имеются меланоциты.
Считается, что клетки Лангерганса играют определенную роль в защитном механизме слизистой оболочки полости рта. Было высказано предположение, что клетки Лангерганса реагируют с антигенами, которые находятся в процессе проникновения в эпителий. Таким образом, инициируется ранний иммунологический ответ, инициирующий, ингибирующий или предотвращающий дальнейшее проникновение антигена в ткань. Предполагается, что клетки Меркеля обладают сенсорной функцией.
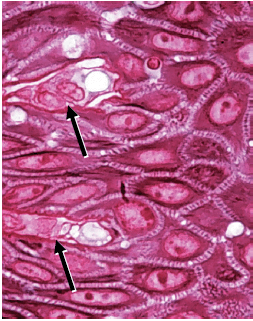
Клетки базального слоя имеют цилиндрическую или кубовидную форму и находятся в контакте с основной мембраной, которая разделяет эпителий и соединительную ткань. Базальные клетки обладают способностью к делению, то есть подвергаются митотическому делению клеток. Клетки, отмеченные стрелками на микрофотографии (рис. 20), находятся в процессе деления. Именно в базальном слое происходит обновление эпителия. Поэтому этот слой также называют stratum germinativum, и его можно считать предшественником клеточного эпителия.
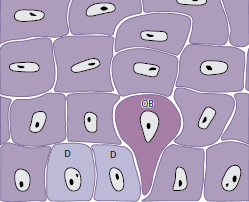
Когда две дочерние клетки (D) были сформированы путем деления клеток, соседняя “более старая” базальная клетка (OB) выталкивается в шиповатый клеточный слой и начинает, как кератиноцит, пересекать эпителий (рис. 21). Кератиноциту требуется примерно 1 месяц, чтобы достичь наружной поверхности эпителия, где он «сбрасывается» из рогового слоя. В течение данного времени число клеток, которые делятся в базальном слое, равно числу клеток, которые «сбрасываются» с поверхности. Таким образом, в нормальных условиях существует равновесие между обновлением клеток и их потерей, так что эпителий сохраняет постоянную толщину.
По мере того как базальная клетка мигрирует через эпителий, она становится уплощенной со стороны длинной оси, параллельной поверхности эпителия.
Базальные клетки находятся непосредственно рядом с соединительной тканью и отделены от нее базальной мембраной, вероятно формируемой базальными клетками. Под световым микроскопом (рис. 22) эта мембрана выглядит как бесструктурная зона шириной примерно 1-2 мкм (стрелки) и положительно реагирует на кислотно-шиффовое окрашивание (PAS). Эта положительная реакция свидетельствует о том, что базальная мембрана содержит углеводы (гликопротеины). Эпителиальные клетки окружены внеклеточным веществом, которое также содержит белково–полисахаридные комплексы. На ультраструктурном уровне базальная мембрана имеет сложный состав.
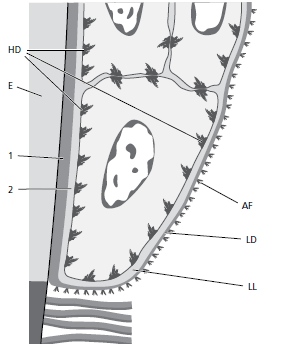
Представлена электронная микрофотография (рис. 23) (увеличение×70000) участка, включающего часть базальной клетки, базальную мембрану и часть прилегающей соединительной ткани.
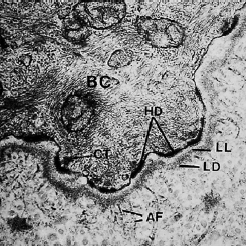
Базальная клетка (BC) занимает верхнюю часть микрофотографии. Непосредственно под базальной клеткой можно увидеть электрон‐светящуюся зону шириной примерно 400 Å, которая называется lamina lucida (LL). Под пластинкой lucida можно наблюдать электронно‐плотную зону примерно такой же толщины. Эта зона называется lamina densa (LD). Из темной пластинки так называемые якорные волокна (AF) веерообразно входят в соединительную ткань. Якорные волокна примерно равны 1 мкм в длину и свободно заканчиваются в соединительной ткани. Базальная мембрана, которая под световым микроскопом выглядит как единое целое, на электронной микрофотографии, по-видимому, состоит из одной пластинки lucida и одной пластинки densa с прилегающими соединительнотканными волокнами (якорными волокнами). Клеточная мембрана эпителиальных клеток, обращенных к пластинке lucida, содержит ряд электронно‐плотных, более толстых зон, появляющихся через различные интервалы вдоль клеточной мембраны. Эти структуры называются hemidesmosomes (HD). Цитоплазматические тонофиламенты (СТ) в клетке сходятся к хемидесмосомам. Хемидесмосомы участвуют в прикреплении эпителия к нижележащей базальной мембране.
На рис. 24 показана область шиповатого слоя в десневом эпителии полости рта. Stratum spinosum состоит из 10-20 слоев относительно крупных клеток, снабженных короткими цитоплазматическими отростками. Цитоплазматические отростки (стрелки) отходят через равные промежутки и дают клеткам шиповатый вид. Вместе с межклеточными белково–углеводными комплексами, сцепление между клетками обеспечивается многочисленными “десмосомами” (парами гемидесмосом), которые располагаются между цитоплазматическими отростками соседних клеток.
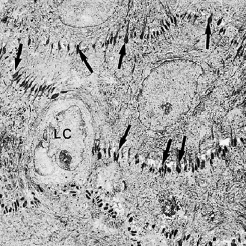
На рис. 25 показана область stratum spinosum на электронной микрофотографии. Темноватые структуры между отдельными эпителиальными клетками представляют собой десмосомы (стрелки). Десмосому можно рассматривать как две гемидесмосомы, обращенные друг к другу.
Наличие большого количества десмосом указывает на то, что сцепление между эпителиальными клетками является прочным. Светлая клетка (LC) в центре микрофотографии не содержит гемидесмосом и поэтому является не кератиноцитом, а скорее “прозрачной клеткой”.
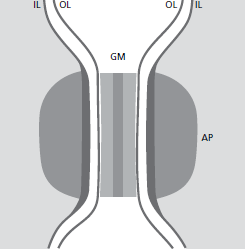
Схематичный рисунок 26, показывающий состав десмосомы. Десмосому можно считать состоящей из двух соседних гемидесмосом, разделенных зоной, содержащей электронно‐плотный гранулированный материал (GМ). Таким образом, десмосома состоит из следующих структурных компонентов: (1) наружные листки (OL) клеточных мембран двух соседних клеток, (2) толстые внутренние листки (IL) клеточных мембран и (3) прикрепляющиеся бляшки (AP), которые представляют собой зернистый и фибриллярный компонент в цитоплазме.
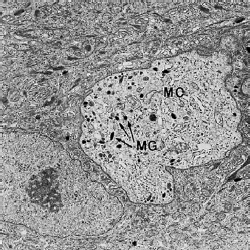
Как упоминалось ранее, эпителий полости рта также содержит меланоциты, которые отвечают за выработку пигмента меланина. Меланоциты присутствуют у лиц с выраженной пигментацией слизистой оболочки полости рта, а также у лиц, у которых невозможно заметить клинические признаки пигментации. На этой электронной микрофотографии меланоцит (МC) присутствует в нижней части шиповатого слоя. В отличие от кератиноцитов, эта клетка содержит гранулы меланина (МG) и не имеет ни тонофиламентов, ни гемидесмосом. Обратите внимание на большое количество тонофиламентов в цитоплазме соседних кератиноцитов.
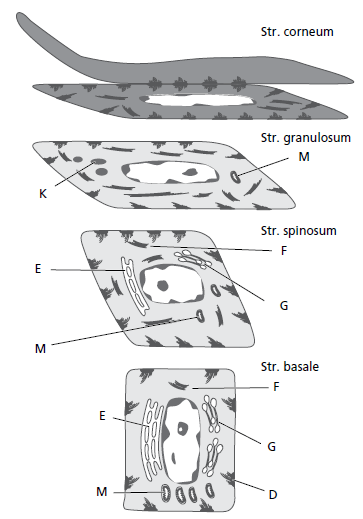
При пересечении эпителия от базального слоя к поверхности эпителия кератиноциты проходят непрерывную дифференциацию и специализацию. Многочисленные изменения, которые происходят во время этого процесса, показаны на этой диаграмме (рис. 28) ороговевшего многослойного плоского эпителия. От базального слоя (stratum basale) к зернистому слою (stratum granulosum) увеличивается как количество тонофиламентов (F) в цитоплазме, так и количество десмосом (D). Напротив, количество органелл, таких как митохондрии (M), пластинки грубого эндоплазматического ретикулума (E) и комплексы Гольджи (G), уменьшается в кератиноцитах на их пути из базального слоя к поверхности. В зернистом слое начинают появляться электронно‐плотные кератогиалиновые тела (К) и скопления гликогенсодержащих гранул. Считается, что такие гранулы связаны с синтезом кератина.
Микрофотография (рис. 29) зернистого и рогового слоев. Гранулы кератогиалина (стрелки) видны в зернистом слое.
Происходит резкий переход клеток из зернистого слоя в роговой. Это указывает на очень внезапную кератинизацию цитоплазмы кератиноцита и его переход в роговые чешуйки. Цитоплазма клеток в роговом слое (SС) заполнена кератином и весь аппарат для синтеза белка и производства энергии, то есть ядро, митохондрии, эндоплазматический ретикулум и комплекс Гольджи, теряется. В паракератинизированном эпителии, однако, клетки рогового слоя содержат остатки ядер. Кератинизация считается процессом дифференцировки, а не дегенерации. Это процесс синтеза белка, который требует энергии и зависит от функциональных клеток, то есть клеток, содержащих ядро и нормальный набор органелл.
Суммарно: кератиноцит подвергается непрерывной дифференцировке на своем пути от базального слоя к поверхности эпителия. Таким образом, как только кератиноцит покинул базальную мембрану, он больше не может делиться, но сохраняет способность к производству белка (тонофиламентов и кератогиалиновых гранул). В зернистом слое кератиноцит лишается своего энергетического и белкового аппарата (вероятно, в результате ферментативного расщепления) и резко превращается в кератиновую клетку, которая через роговой слой, «сбрасывается» с поверхности эпителия.
На рис. 30 показана часть эпителия альвеолярной (выстилающей) слизистой оболочки. В отличие от эпителия десны, выстилающая слизистая оболочка не имеет рогового слоя. Отметим, что клетки, содержащие ядра, можно идентифицировать во всех слоях, от базального слоя до поверхности эпителия.
Зубо-десневой эпителий
Ткани зубодесневой области достигают своей зрелости в сочетании с прорезыванием зубов (рис. 31).
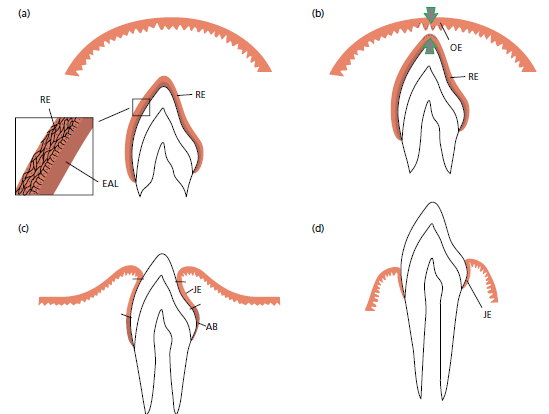
а — когда эмаль зуба полностью сформирована, клетки, продуцирующие эмаль (амелобласты) уменьшаются в высоту, формируют структуры базальной пластинки и образуют вместе с клетками из наружного эпителия эмали так называемый редуцированный зубной эпителий (RE). Базальная пластинка (эпителиального прикрепления пластинки[EAL]) находится в непосредственном контакте с эмалью. Контакт между этой пластинкой и эпителиальными клетками поддерживается гемидесмосомами. Редуцированный эпителий эмали окружает коронку зуба с момента правильной минерализации эмали пока зуб не начнет прорезываться.
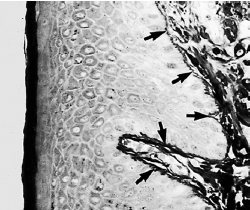
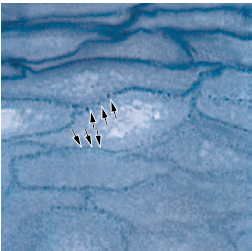
b — по мере приближения прорезывающегося зуба к эпителию полости рта, клетки наружного слоя редуцированного зубного эпителия (RE), а также клетки
базального слоя эпителия полости рта (ОE) проявляют повышенную митотическую активность (стрелки) и начинают мигрировать в нижележащую соединительную ткань. Мигрирующий эпителий образует эпителиальную массу между оральным эпителием и редуцированным зубным эпителием, так что зуб может прорезаться без кровотечения. Прежние амелобласты не делятся.
с — когда зуб уже частично в ротовой полости, большие участки режущего края эмали покрыты соединительным эпителием (JE), содержащим только несколько слоев клеток. Пришеечная область эмали, однако, все еще покрыта амелобластами (АB) и наружными клетками редуцированного зубного эпителия.
d — во время более поздних фаз прорезывания зуба все клетки редуцированного эпителия эмали заменены соединительным эпителием (JE). Этот эпителий неразрывно связан с эпителием полости рта и обеспечивает прикрепление между зубом и десной. Если свободная десна иссекается после полного прорезывания зуба, то во время заживления образуется новый соединительный эпителий, неотличимый от того, который обнаруживается после прорезывания зуба. Тот факт, что этот новый соединительный эпителий развился из эпителия полости рта, указывает на то, что клетки этого эпителия обладают способностью дифференцироваться в клетки соединительного эпителия.
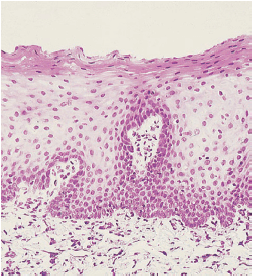
На рис. 32 представлен гистологический разрез через границу между зубом и десной, то есть зубодесневую область.
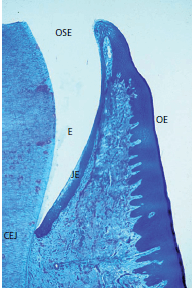
Эмаль (Е) находится слева. Справа соединительный эпителия (JE), бороздчатый эпителий полости рта (ОSE), и ротовой эпителий (ОE). Эпителий борозды покрывает неглубокую десневую бороздку, , расположенную между эмалью и краем свободной десны. Соединительный эпителий морфологически отличается от бороздчатого эпителия и орального эпителия, в то время как последние два структурно очень похожи. Хотя отдельные вариации могут иметь место, соединительный эпителий обычно наиболее широкий в своей корональной части (около 15-20 слоев клеток), но становится тоньше (3-4 клетки) по направлению к цементо-эмалевому соединению (CEJ). Граница между соединительным эпителием и подлежащей соединительной тканью не имеет эпителиальных ретикулоцитов, за исключением случаев воспаления.
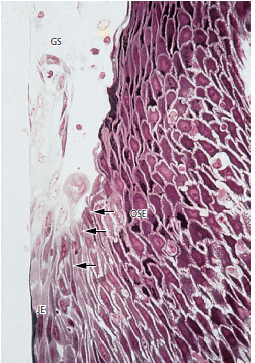
Соединительный эпителий (рис. 33) имеет свободную поверхность в нижней части десневой борозды (GS). Как и эпителий борозды и ротовой эпителий, соединительный эпителий непрерывно обновляется за счет деления клеток в базальном слое. Клетки мигрируют к основанию десневой борозды, откуда они слущиваются. Граница между соединительным эпителием (JE) и ротовым бороздчатым эпителием (OSE) обозначается стрелками. Клетки ротового бороздчатого эпителия имеют кубовидную форму, и поверхность этого эпителия не кератинизирована.
На рис. 34 показаны различные характеристики соединительного эпителия.
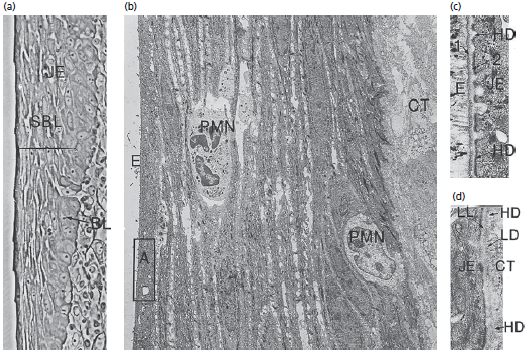
Как видно на a, клетки соединительного эпителия (JE) расположены в одном базальном слое (BL) и нескольких супрабазальных слоях (SBL), b- показано, что базальные клетки, а также супрабазальные клетки уплощены по длинной оси, параллельной поверхности зуба. (СТ, соединительная ткань; E, эмалевое пространство.)
Существуют четкие различия между эпителием борозды, оральным эпителием и соединительным эпителием:
- Размер клеток в соединительном эпителии относительно объема ткани больше, чем в ротовом
эпителии. - Межклеточное пространство в соединительном эпителии является, по отношению к объему ткани, сравнительно более широким, чем в ротовом эпителии.
- Количество десмосом в соединительном эпителии меньше, чем в эпителии полости рта.
Обратите внимание на сравнительно широкие межклеточные пространства между продолговатыми клетками соединительного эпителия и наличие двух нейтрофильных гранулоцитов (PMN), которые пересекают эпителий.
Область (а) это то, что показано на схеме с под увеличением. Видно, что базальные клетки соединительного эпителия не находятся в непосредственном контакте с эмалью (Е). Между эмалью и эпителием (JE) можно увидеть одну электронно‐темную зону (1) и одну электронно‐светлую зону (2). Электронно‐светлая зона находится в контакте с клетками соединительного эпителия ( JE). Эти две зоны имеют структуру очень похожую на структуру lamina densa (LD) и lamina lucida (LL) в области базальной мембраны. Кроме того, как видно на d, клеточная мембрана соединительных эпителиоцитов содержит гемидесмосомы (HD) по направлению к эмали и соединительной ткани. Таким образом, граница раздела между эмалью и соединительным эпителием аналогична границе раздела между эпителием и соединительной тканью.
Представлен схематический рисунок 35 наиболее апикально расположенной клетки в соединительном эпителии. Эмаль (Е) изображена слева. Можно видеть, что электронно-темная зона (1) между соединительным эпителием и эмалью может рассматриваться как продолжение lamina densa (LD) в базальной мембране соединительно тканной стороны. Аналогично, электронно-светлую зону (2) можно рассматривать как продолжение lamina lucida (LL). Следует, однако, отметить, что в отличие от вида эпителио–соединительной ткани, здесь нет никаких якорных волокон (АF), прикрепленных к пластинчатой структуре densa (1) рядом с эмалью. С другой стороны, подобно базальным клеткам, примыкающим к базальной мембране (на соединительнотканный стороне), клетки соединительного эпителия, обращенные к структуре, напоминающей lamina lucida (2), содержат гемидесмосомы (HD). Таким образом, взаимодействие между соединительным эпителием и эмалью структурно очень похож на взаимодействие эпителия и соединительной ткани, что означает, что соединительный эпителий не только контактирует с эмалью, но и фактически физически прикреплен к зубу через гемидесмосомы.
Собственная пластинка
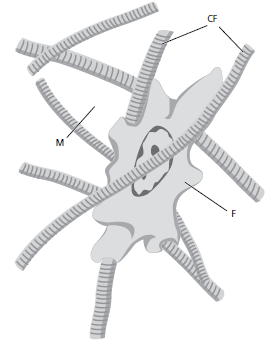
Преобладающим тканевым компонентом десны является соединительная ткань (lamina propria). Крупными компонентами соединительной ткани являются коллагеновые волокна (около 60% объема соединительной ткани), фибробласты (около 5%), сосуды и нервы (около 35%), которые встроены в аморфное основное вещество (матрикс).
На рис. 36 показан фибробласт (F), находящийся в сети соединительнотканных волокон (CF). Промежуточное пространство заполнено матрицей (М), которая образует “среду” для клетки.
Продолжение статьи «Строение пародонта: анатомия и гистология тканей пародонта. Часть 2».
Перевод материала «Anatomy of Periodontal Tissues» Jan Lindhe, Thorkild Karring, Maurício Araújo выполнен специально для проекта «Современная пародонтология».