Кость — это сложный орган, состоящий из множества специализированных тканей (костная, надкостница / эндост и костный мозг), которые действуют синергетически и выполняют множество функций (рис. ниже). Ее состав позволяет костной ткани: (1) выдерживать нагрузку, (2) защищать высокочувствительные органы от внешних сил и (3) участвовать в качестве хранилища клеток и минералов, которые способствуют системному гомеостазу тела.
Таким образом, концепция «кость как живой орган» объединяет структурно-динамическую природу кости с ее способностью управлять множеством механических и метаболических функций с важными локальными и системными последствиями. На эту систему оказывают влияние множество факторов (например, биохимические, гормональные, клеточные, биомеханические), которые в совокупности определяют ее качество (Ammann & Rizzoli 2003; Marotti & Palumbo 2007; Bonewald & Johnson 2008; Ma et al. 2008). Цель этого материала — предоставить фундаментальные знания о развитии, структуре, функции, заживлении и гомеостазе костной ткани.
Костный орган включает в себя ряд сложно организованных тканей, которые в состоянии здоровья взаимодействуют друг с другом для выполнения ряда функций. Он служит источником стволовых клеток и хранилищем минералов и других питательных веществ; защищает ряд чувствительных органов; а также действует как механический датчик, который адаптируется к окружающей среде и индивидуальным требованиям. На этом рисунке выделены три основные ткани и клетки, которые участвуют в выполнении основных функций и поддержании структуры и функции кости как органа (DFCT- плотная волокнистая соединительная ткань; LFCT- рыхлая волокнистая соединительная ткань).
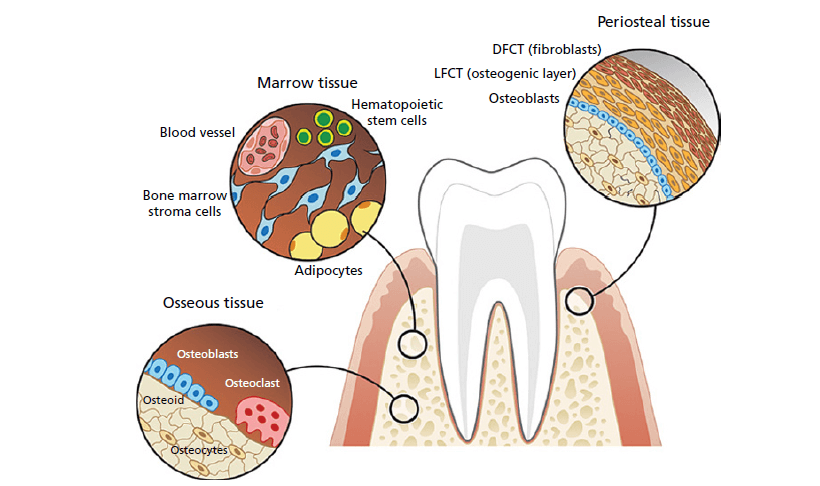
Развитие
Во время эмбриогенеза скелет формируется путем прямого или косвенного процесса окостенения. В случае с нижней челюстью, верхней челюстью, черепом и ключицей мезенхимальные клетки-предшественники конденсируются и подвергаются прямой дифференцировке в остеобласты, процесс, известный как внутримембранный остеогенез.
Напротив, в мыщелке нижней челюсти, длинных костях и позвонках первоначально образуется модель хряща, который служит зачатком, постепенно заменяющимся костью. Хрящ-зависимый процесс формирования и роста костей известен как эндохондральный остеогенез (Ranly 2000) (рис. ниже).
Внутримембранный остеогенез
Во время внутримембранного остеогенеза центр окостенения развивается за счет мезенхимальной конденсации. По мере развития и созревания богатого коллагеном внеклеточного матрикса клетки остеопрогенератора подвергаются дальнейшей остеобластической дифференцировке. На наружных поверхностях очага окостенения над слоем остеобластов образуется фиброзная надкостница. По мере того как новые остеобласты формируются с нижней стороны надкостницы, происходит аппозиционный рост. Субпопуляция остеобластов внедряется в минерализующий матрикс и дает начало лакуно-каналикулярной сети остеоцитов. В пределах черепно-лицевого комплекса большинство костей развивается и растет благодаря этому механизму.
Рост эндохондральной кости
Во время эндохондрального остеогенеза кости развиваются за счет образования хрящевой матрицы (гиалиновой модели хряща), которая минерализуется, а затем рассасывается остеокластами и заменяется костью, которая впоследствии откладывается.
Этот процесс начинается на третьем месяце беременности. Процесс эндохондрального окостенения приводит к образованию первичных и вторичных центров окостенения, которые разделены хрящевой структурой, известной как пластинка роста.
После образования первичного очага окостенения костное образование распространяется к обоим концам кости от центра ствола. Клетки хряща на передних краях окостенения отмирают.
Остеобласты покрывают хрящевые трабекулы ткаными губчатыми костями. За наступающим фронтом окостенения остеокласты поглощают губчатую кость и увеличивают первичную полость костного мозга. Надкостничный воротник утолщается и расширяется к эпифизам, чтобы компенсировать продолжающееся углубление первичной полости.
Процессы остеогенеза и резорбции происходят во всех направлениях. Пространства между трабекулами заполняются тканью костного мозга. По мере ремоделирования нового костного матрикса остеокласты способствуют образованию первичных мозговых полостей, которые быстро заполняются кроветворной тканью костного мозга. Фиброзная неминерализованная выстилка костномозговой полости — это эндост. Остеобласты образуются в эндосте и начинают формирование эндостальной кости. Аппозиционный рост эндостальной кости тщательно регулируется, чтобы предотвратить закрытие первичных полостей костного мозга и разрушение костного мозга.
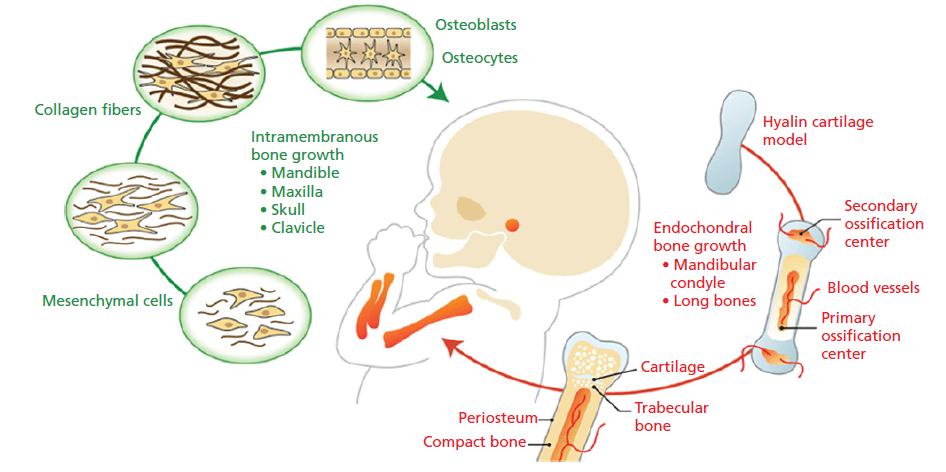
Развитие костей. Существует два типа процессов, участвующих в развитии кости: внутримембранное окостенение (зеленая стрелка) и эндохондральное окостенение (оранжевая стрелка). Они в первую очередь отличаются наличием хрящевого шаблона во время роста эндохондральной кости. Во время внутримембранного остеогенеза центр окостенения развивается за счет мезенхимальной конденсации. По мере развития и созревания богатого коллагеном внеклеточного матрикса клетки остеопрогенератора подвергаются дальнейшей остеобластической дифференцировке. Субпопуляция остеобластов внедряется в минерализующий матрикс и дает начало лакуно-каналикулярной сети остеоцитов. В пределах черепно-лицевого комплекса большинство костей развивается и растет благодаря этому механизму. С другой стороны, длинные кости внутри скелета и мыщелкового отростка нижней челюсти первоначально развиваются за счет образования модели хрящевой ткани, который минерализуется, а затем рассасывается остеокластами и заменяется костью. Процесс роста эндохондральной кости приводит к образованию первичных и вторичных центров окостенения, которые разделены хрящевой структурой, известной как пластинка роста. По мере того как кость развивается и созревает в результате этих двух процессов, структурно различные участки компактной кости и трабекулярной кости формируются и поддерживаются посредством сходных механизмов костного ремоделирования.
Костная ткань. Состав
Костная ткань
Костная ткань — это специализированная соединительная ткань, состоящая из органических и неорганических элементов, которая минерализуется и наполняется высокоспециализированными клетками, регулирующими ее стабильность (рис. ниже).
Матрикс
Органический матрикскости составляет приблизительно 30-35% от общей массы кости и состоит на 90% из коллагена I типа и на 10% из неколлагеновых белков, протеогликанов, гликопротеинов, углеводов и липидов.
Органический матрикс синтезируется остеобластами и, хотя он еще неминерализован, известен как остеоид. Внутри коллагеновых волокон происходит зарождение минералов, когда ионы кальция и фосфата откладываются и в конечном итоге образуют кристаллы гидроксиапатита. Неколлагеновые белки на поверхности коллагеновых волокон способствуют распространению минерала и полной минерализации матрикса.
Неорганические компоненты
Гидратированный кальций и фосфат в форме кристаллов гидроксиапатита [3Ca3 (PO4) 2 (OH) 2] являются основным неорганическим компонентом костного матрикса. Минерализация видна при электронной микроскопии как яркий сигнал (рис. ниже). Конкретные элементы внутри минерала могут быть дополнительно идентифицированы с помощью энергодисперсионной рентгеновской спектроскопии (ЭДС). На рис.ниже характерные пики кальция и фосфора значительно выражены в кости, как и ожидалось из-за их высокого содержания в кристаллах гидроксиапатита.
Органические компоненты
Изначально кость представляет собой чисто органический матрикс, богатый коллагеном, а также другими неколлагеновыми молекулами (рис. ниже). Химический анализ кости с помощью Рамановской спектроскопии четко выделяет этот органический аналог в матрице. Переход от чисто органической матрицы к минерализованной матрице четко показан на микрофотографии просвечивающего электронного микроскопа на рис. ниже, когда остеоцит внедряется в минерализованную зрелую матрицу. По мере созревания матрикса минералнуклеация и размножение опосредуются органическими компонентами внеклеточного матрикса. На рис. показана агрегация кристаллов минералов, образующих круговые структуры. По мере того как минерал распространяется вдоль фибрилл коллагена, формируется четкий фронт минерализации и четко разграничивает переход между остеоидной областью и зрелой костью.
Минерализация
Инициация минерализации внутри остеоида обычно происходит в течение нескольких дней после укладки ионов кальция и фосфата, но созревание завершается за счет роста кристаллов гидроксиапатита в течение нескольких месяцев и по мере синтеза новой матрицы (рис. ниже). Помимо обеспечения прочности и жесткости для сопротивления нагрузкам и защиты высокочувствительных органов, минерализация остеоида позволяет накапливать минералы, способствующие системному гомеостазу.
Клетки
Внутри кости можно идентифицировать различные клеточные компоненты. Отдельные клеточные популяции включают остеогенные клетки-предшественники, остеобласты, остеокласты, остеоциты и кроветворные элементы костного мозга. Мы сосредоточимся на трех основных функциональных клетках, которые в конечном счете отвечают за правильный гомеостаз скелета.
Остеобласты (рис. ниже)
Остеобласты являются первичными клетками, ответственными за формирование кости; они синтезируют компоненты органического внеклеточного матрикса (ЭКМ) и контролируют минерализацию матрикса (Рис. ниже). Остеобласты располагаются на костных поверхностях, демонстрируя активное отложение матрикса, и в конечном итоге могут дифференцироваться в два различных типа клеток: клетки костной выстилки и остеоциты. Клетки костной выстилки — это удлиненные клетки, которые покрывают поверхность костной ткани и не проявляют никакой синтетической активности. Остеобласты являются полностью дифференцированными клетками и не обладают способностью к миграции и пролиферации. Таким образом, для формирования кости в данном участке недифференцированные мезенхимальные клетки-предшественники, движимые экспрессией гена, известного как indian hedgehog («индийский еж»), а затем RUNX2, и остеопрогенерирующие клетки мигрируют в этот участок и пролиферируют, превращаясь в остеобласты (рис. ниже). Определенные клетки остеопрогенератора присутствуют в костном мозге, в эндосте и в надкостнице, которая покрывает поверхность кости. Такие клетки обладают врожденной способностью к пролиферации и дифференцировке в остеобласты. Дифференцировка и развитие остеобластов из остеопрогенерирующих клеток зависят от высвобождения остеоиндуктивных или остеопро‐моторных факторов роста (ОмФ), таких как костные морфогенетические белки (КМгБ), и других факторов роста, таких как инсулиноподобный фактор роста (ИФП), тромбоцитарный фактор роста(ТБФР) и фактор роста фибробластов (ФРФ).
Остеоциты (рис. ниже)
Остеоциты‐это звездчатые клетки, которые встроены в минерализованный костный матрикс в пространствах, известных как лакуны (рис. ниже). Они формируют сеть цитоплазматических отростков, известных как дендриты (рис.ниже).
Эти цитоплазматические проекции остеоцитов проходят через цилиндрические заключенные компартменты, обычно называемые канальцами (Bonewald 2007).). Они распространяются на различные участки и контактируют с кровеносными сосудами и другими остеоцитами (рис.ниже). Таким образом, сеть остеоцитов является внеклеточным и внутриклеточным каналом связи, чувствительным на мембранном уровне к напряжению , вызванному потоком жидкости в канальцевом пространстве в результате механических раздражителей и деформации кости. Остеоциты переводят механические сигналы в биохимические медиаторы, которые помогают в регуляции анаболических и катаболических событий в костной ткани. Такое расположение позволяет остеоцитам (1) участвовать в регуляции кальциевого гомеостаза крови и (2) ощущать механическую нагрузку и передавать эту информацию другим клеткам внутри кости для дальнейшего управления функцией остеобластов и остеокластов (Burger et al. 1995; Marotti 2000). Различные костные заболевания и нарушения влияют на расположение лакуноканаликулярной системы остеоцитов, вызывая значительные изменения в этой важной клеточной сети (рис. ниже).
Остеокласты
Активность костеобразования последовательно связана с резорбцией кости, которая инициируется и поддерживается остеокластами. Эти клетки обладают способностью развиваться и прилипать к костному матриксу, а затем выделять кислые и литические ферменты, которые разрушают и расщепляют минеральные и органические компоненты кости и кальцинированного хряща (рис. ниже). Процесс деградации матрикса приводит к образованию специализированного внеклеточного компартмента, известного как лакуна Хаушипа (Rodan 1992;Vaananen & Laitala‐Leinonen 2008). Остеокласты — это специализированные многоядерные клетки, происходящие из моноцитарно-макрофагальной кроветворной линии. Процесс дифференцировки первоначально обусловлен экспрессией транскрипционного фактора PU‐1. Макрофагальный колониестимулирующий фактор (м‐КСФ) вовлекает остеокласты в путь дифференцировки и способствует их пролиферации и экспрессии RANKL. На этой стадии стромальные клетки, экспрессирующие RANKL, взаимодействуют с преостеокластами и далее связывают их с дифференцировкой по линии остеокластов (рис. ниже).
Надкостничная ткань
Надкостница-это волокнистая оболочка, которая выстилает наружную поверхность стержня длинной кости (диафиз), но не суставные поверхности. Эндост выстилает внутреннюю поверхность всех костей. Надкостница состоит из плотной нерегулярной соединительной ткани. Надкостница делится на плотный, волокнистый, сосудистый слой (“фиброзный слой”) и внутренний, более рыхлый, соединительнотканный внутренний слой (“остеогенный слой”) (см. рис.). Фиброзный слой в основном состоит из фибробластов, в то время как внутренний слой содержит остеопрогенерирующие клетки. Остеобласты, полученные из “остеогенного слоя”, ответственны за увеличение ширины длинных костей и общего размера других типов костей. В случае перелома клетки-предшественники из надкостницы дифференцируются в остеобласты и хондробласты, которые играют важную роль в процессе стабилизации раны.
В отличие от костной ткани, надкостница имеет ноцицептивные нервные окончания, что делает ее очень чувствительной к манипуляциям. Он также обеспечивает прохождение лимфатических и кровеносных сосудов в кости и из них, обеспечивая питание. Надкостница прикрепляет сухожилия и связки к кости коллагеновыми волокнами в “остеогенном слое”, называемом волокнами Шарпи, которые простираются до наружных окружных и интерстициальных пластинок. Она также обеспечивает крепление для мышц и сухожилий.
Костный мозг
Костный мозг состоит из островков кроветворной ткани, стромальных клеток и жировых клеток, окруженных сосудистыми синусами, вкрапленными в сетку трабекулярной кости (см. рис.). Костный мозг является основным кроветворным органом, первичной лимфоидной тканью (ответственной за производство эритроцитов, гранулоцитов, моноцитов, лимфоцитов и тромбоцитов) и важным источником стволовых клеток.
Типы
Существует два типа костного мозга: красный, который состоит в основном из кроветворной ткани, и желтый, который в основном состоит из адипоцитов. Эритроциты, лейкоциты и тромбоциты возникают в красном мозге. Оба типа костного мозга содержат многочисленные кровеносные сосуды и капилляры. При рождении весь костный мозг красный. С возрастом все больше и больше его превращается в желтый тип; только около половины взрослого костного мозга красного цвета. В случаях тяжелой кровопотери организм может преобразовать желтый костный мозг обратно в красный, чтобы увеличить производство клеток крови.
Клетки
Основа костного мозга не участвует непосредственно в первичной функции кроветворения. Однако он играет косвенную роль, обеспечивая идеальное кроветворное микроокружение.
Например, он вырабатывает колониестимулирующие факторы, которые оказывают значительное влияние на кроветворение. Клетками, составляющими основу костного мозга, являются:
• Фибробласты
• Макрофаги
• Адипоциты
• Остеобласты
• Остеокласты
• Эндотелиальные клетки.
Стволовая клетка
Основа костного мозга – это мезенхимальные стволовые клетки (МСК), также называемые стромальными клетками костного мозга. Это мультипотентные стволовые клетки, которые могут дифференцироваться в различные типы клеток. Было показано, что МСК дифференцируются in vitro (полученный в искусственных условиях; в искусственном окружении) или in vivo (в естественных условиях; на живом организме) в остеобласты, хондроциты, миоциты, адипоциты и бета‐панкреатические островковые клетки.
МСК также могут трансдифференцироваться в нейрональные клетки. Кроме того, костный мозг содержит гемопоэтические стволовые клетки, которые дают начало трем классам клеток крови, которые находятся в кровообращении: лейкоциты, эритроциты и тромбоциты.
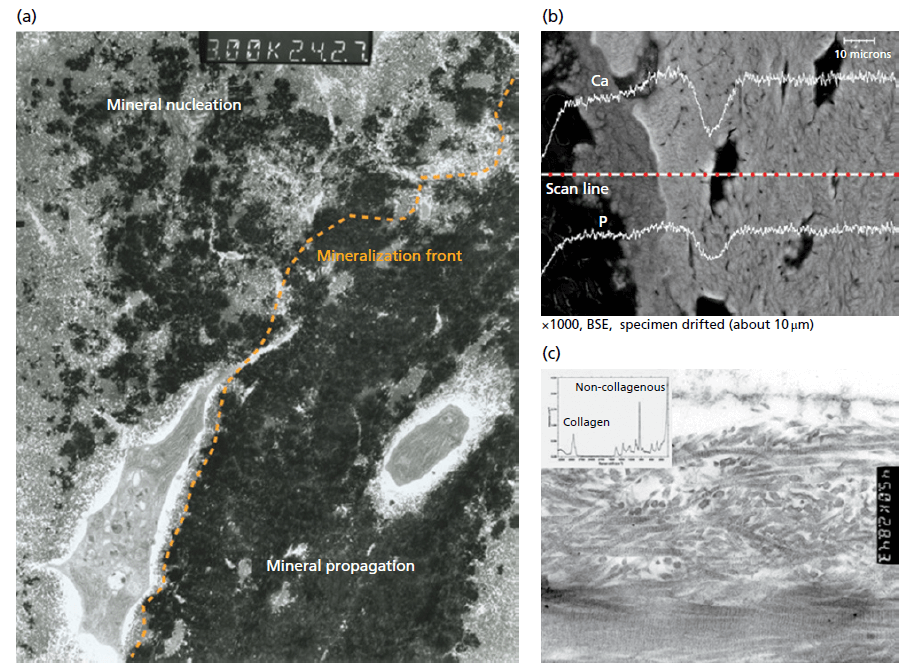
Костный матрикс. Внеклеточный матрикс в кости особенно богат по сравнению с его клеточным аналогом. а) костный матрикс обладает уникальной способностью к минерализации: этот процесс требует поддержки органических компонентов и помощи высокоспециализированных клеток. b) кальций и фосфор присутствуют в виде кристаллов гидроксиапатита. Эти кристаллы, как правило, следуют за органическим каркасом в костном матриксе. Оранжевая пунктирная линия представляет собой линейное сканирование, которое подчеркивает высокое содержание кальция и фосфора в зрелой кости, как показал рентгеновский спектроскопический анализ. с) коллагеновые волокна, а также неколлангенные белки в изобилии присутствуют в матриксе и часто лежат в таком направлении, как это показано на спектроскопии.
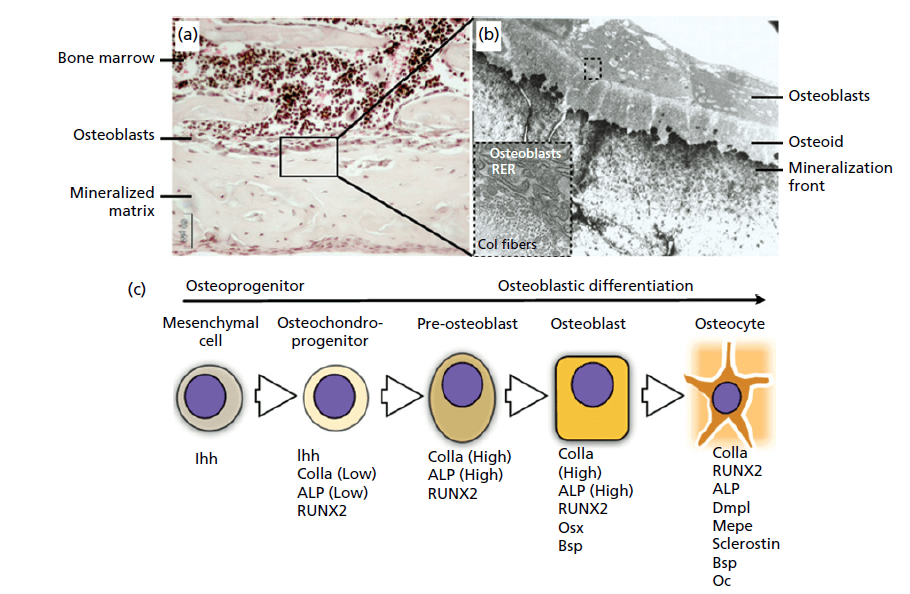
Остеобласт. Остеобласты происходят из клеток-остеопрогениторов костного мозга и отвечают за синтез незрелого костного матрикса, известного как остеоид. (а) Группа остеобластов, выстилающих зрелую кость, которая содержит клетки, встроенные в минерализованный матрикс. (b) Группы остеобластных клеток четко видны с помощью электронной микроскопии (ЭМ). Выраженный грубый эндоплазматический ретикулум (ГЭР) и аппарат Гольджи внутри этих клеток отражают их высокую метаболическую активность. (c) На схеме показана дифференцировка остеопрогениторной клетки до зрелого дифференцированного остеоцита.

Остеоциты. Остеоцит можно определить как двигатель процесса ремоделирования внутри костной ткани. (а) по мере синтеза костного матрикса ряд остеобластов внедряется в остеоид, который затем минерализуется, они в свою очередь находятся в зрелом матриксе уже в виде остеоцитов, как показано на этой сканирующей электронной микрофотографии, обработанной осмием, чтобы обеспечить визуализацию клетки. (b) остеоциты находятся в четко определенном пространстве в кости, известном как остеоцитарная лакуна. с) электронная микрофотография дендрита в канальцах, показывающая пространство, которое заполнено жидкостью; напряжение сдвига стимулирует поверхность клеточной мембраны остеоцита. Эта уникальная биологическая архитектурная характеристика остеоцита и лакуно-каналикулярной сети представляют собой основу, позволяющую преобразовывать механические стимулы в биохимические сигналы, необходимые для гомеостаза кости. (d и e) Микрофотография лакуно-каналикулярной сети позволяет визуализировать степень связи между остеоцитами и регулярно расположенными каналикулярными структурами.
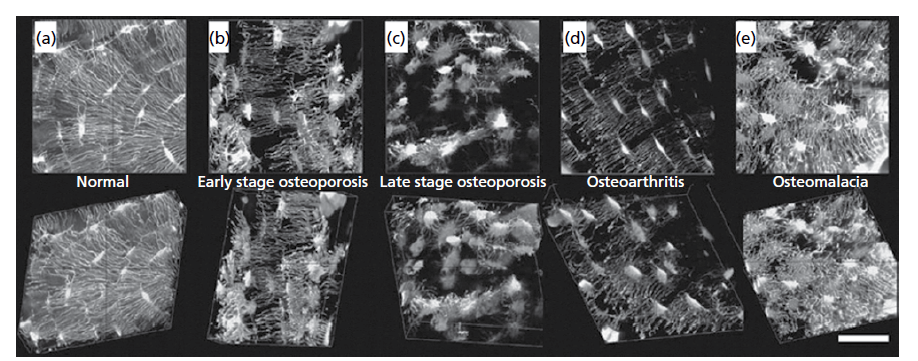
Остеоциты. Лакуно-каналикулярная система при патологии а) в здоровой кости имеется высокая плотность остеоцитов по всему зрелому матриксу и имеется высокая клеточная взаимосвязь. При патологии система значительно нарушается, что приводит к важным функциональным изменениям. (b, c) при остеопорозе наблюдается изменение плотности остеоцитов и явное снижение клеточной взаимосвязанности. d) при остеоартрите изменяется сама канальцевая система, но без существенных лакунарных изменений. е) при остеомаляции вся лакуно-каналикулярная система остеоцитов, по-видимому, нарушается из-за измененного характера минерализации (Источник: Knothe Tate et al. 2004).
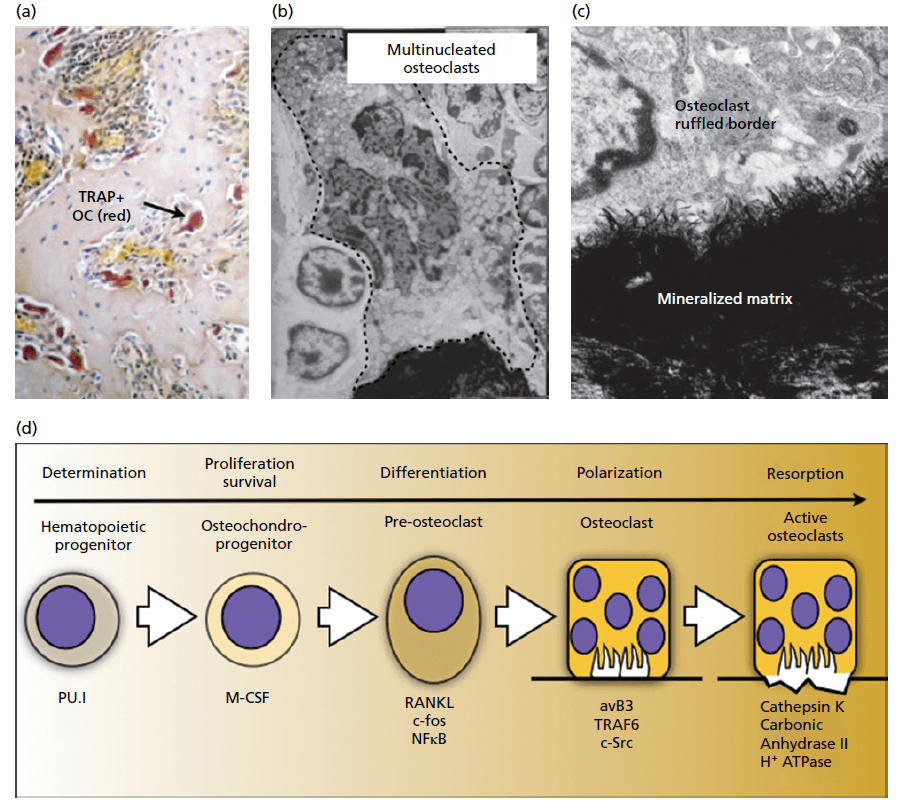
Остеокласты. (а) гистологически остеокласты представляют собой многоядерные клетки, прикрепленные к костному матриксу с помощью специальных структур, обозначаемых как тартраторезистентное кислотное фосфатазное пятно (TRAP) (стрелка). (ОС, остеокласт). (б) электронная микрофотография многоядерного остеокласта, прикрепленного к минерализованному костному матриксу, очерчена пунктирной линией. в) гофрированная кайма на резорбирующем конце клеток. (d) остеокласты происходят из клеток макрофагальной/моноцитарной линии и представляют собой костные резорбирующие единицы. Показаны ключевые молекулы, участвующие в ранних этапах дифференцировки гематопоэтического предшественника до зрелого функционального остеокласта.
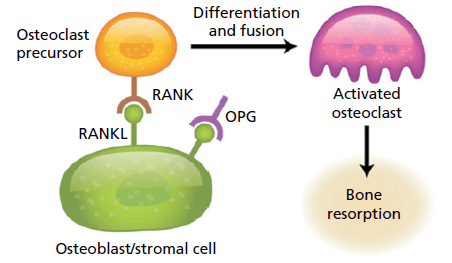
Связь остеообразования и резорбции. Эти процессы тесно связаны между собой. Остеобластные/стромальные клетки обеспечивают остеокластогенное микроокружение (обозначаемое как RANKL) предшественнику остеокластов, вызывая их дальнейшую дифференцировку и слияние, а также приводя к образованию многоядерных и активных остеокластов. Этот процесс модулируется ингибиторами этих взаимодействий, такими как остеопротегерин (ОПГ). Кроме того, образование кости остеобластами зависит от предшествующей резорбции остеокластами.
Перевод материала «Bone as a Living Organ» Hector F. Rios, Jill D. Bashutski, William V. Giannobile выполнен специально для проекта «Современная пародонтология».