Данный материал является продолжением статьи «Строение пародонта: анатомия и гистология тканей пародонта. Часть 1».
Различные типы клеток, присутствующих в соединительной ткани: (1) фибробласты, (2) тучные клетки, (3) макрофаги и (4) воспалительные клетки.
На рис. 37 показан фибробласт является преобладающей клеткой соединительной ткани (65% от общей популяции клеток). Фибробласт участвует в производстве различных типов волокон, обнаруженных в соединительной ткани, но также играет важную роль в синтезе соединительнотканного матрикса. Фибробласт представляет собой веретенообразную или звездчатую клетку с овальным ядром, содержащим одно или несколько ядрышек. Часть фибробластов показана на увеличении. Цитоплазма содержит хорошо развитый гранулярный эндоплазматический ретикулум (Е) с рибосомами. Комплекс Гольджи (G) обычно имеет значительные размеры, а митохондрии (M) крупные и многочисленные. Кроме того, цитоплазма содержит много тонких тонофиламентов (F). Рядом с клеточной мембраной, по всей периферии клетки, можно увидеть большое количество везикул (V).
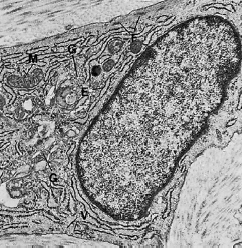
Тучная клетка отвечает за производство определенных компонентов матрицы. Эта клетка также производит вазоактивные вещества, которые могут влиять на функцию микрососудистой системы и контролировать поток крови через ткань. На рис. 38 клетка показана при электронно-микроскопическом увеличении. Цитоплазма характеризуется наличием большого количества везикул (V) различной величины. Эти везикулы содержат биологически активные вещества, такие как протеолитические ферменты, гистамин и гепарин. Комплекс Гольджи (G) хорошо развит, в то время как зернистые структуры эндоплазматического ретикулума редки. Большое количество небольших цитоплазматических отростков, то есть микроворсинок (МV), можно увидеть по периферии клетки.
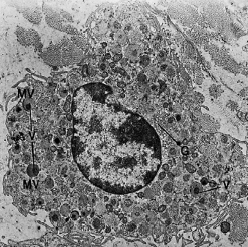
Макрофаг имеет ряд различных фагоцитарных и синтетических функций в тканях. Макрофаг показан при электронно — микроскопическом увеличении на рис. 39. Ядро характеризуется многочисленными инвагинациями различной величины. По периферии ядра видна зона электронно‐ плотных конденсаций хроматина. Комплекс Гольджи (G) хорошо развит, и в цитоплазме присутствуют многочисленные пузырьки (V) различного размера. Гранулярный эндоплазматический ретикулум (Е) скуден, но определенное количество свободных рибосом (R) равномерно распределено в цитоплазме. Остатки фагоцитированного материала часто
обнаруживается в лизосомальных везикулах: фагосомах (РН). На периферии клетки можно увидеть большое количество микроворсинок различного размера. Макрофаги особенно многочисленны в воспаленной ткани. Они образуются из циркулирующих в крови моноцитов, которые мигрируют в ткани.
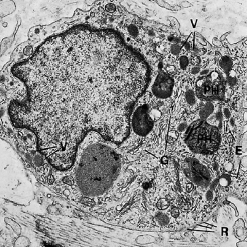
Кроме фибробластов, тучных клеток и макрофагов, соединительная ткань также содержит воспалительные клетки различных типов, например нейтрофильные гранулоциты, лимфоциты и плазматические клетки.
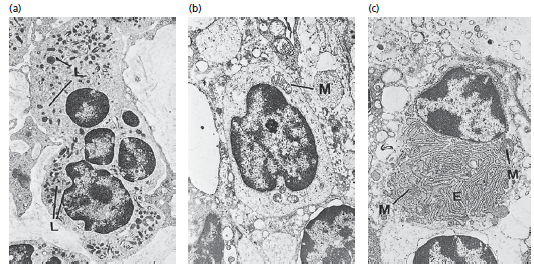
на рис. 40: a — нейтрофильные гранулоциты, также называемые полиморфными-ядерные лейкоциты, имеют характерный внешний вид. Ядро дольчатое, а в цитоплазме обнаружены многочисленные лизосомы (L), содержащие лизосомальные ферменты.
b — лимфоциты характеризуются овальным или сферическим ядром, содержащим локализованные участки электронно‐плотного хроматина. Узкая граница цитоплазмы, окружающая ядро, содержит многочисленные свободные рибосомы, несколько митохондрий (М) и, в локализованных областях, эндоплазматический ретикулум с фиксированными рибосомами. Лизосомы также присутствуют в цитоплазме.
с — плазматические клетки содержат эксцентрично расположенное сферическое ядро с радиально развернутым электронно‐плотным хроматином. Эндоплазматический ретикулум (Е) с многочисленными рибосомами обнаруживается беспорядочно распределенным в цитоплазме. Кроме того, цитоплазма содержит многочисленные митохондрии (М) и хорошо развитый комплекс Гольджи.
Волокна соединительной ткани производятся фибробластами и могут быть разделены на: (1) коллагеновые волокна, (2) ретикулярные волокна, (3) окситалановые волокна и (4) эластические волокна.
Коллагеновые волокна преобладают в десневой соединительной ткани и составляют наиболее существенные компоненты пародонта. На электронной микрофотографии показаны поперечные и продольные сечения коллагеновых волокон. Коллагеновые волокна имеют характерную поперечную исчерченность с периодичностью 700 Å между отдельными темными полосами (рис. 41).
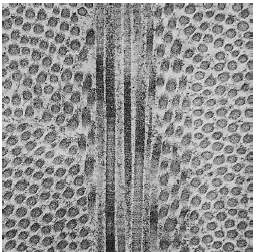
На рис. 42 показаны некоторые важные особенности синтеза и состава коллагеновых волокон, продуцируемых фибробластами (F). Самая маленькая структурная единица тропоколлагена. Тропоколлаген (ТС), который виден в верхней часть рисунка, примерно 3000 Å длиной и имеет диаметр 15 Å. Он состоит из трех полипептидных цепей, переплетенных в спираль. Каждая цепь содержит около 1000 аминокислот. Треть из них -глицин и около 20% — пролин и гидроксипролин, причем последний содержится почти исключительно в коллагене. Синтез тропоколлагена происходит внутри фибробластов, из которых молекула тропоколлагена секретируется во внеклеточное пространство. Таким образом, полимеризация молекул тропоколлагена в коллагеновые волокна происходит внеклеточно. Сначала, структуры тропоколлагена агрегируются в продольном направлении с образованием протофибрилл (PF), которые впоследствии «собираются» латерально параллельно коллагеновым фибриллам (CFR), причем молекулы тропоколлагена перекрываются примерно на 25% их длины. В связи с тем, что после окрашивания в местах соприкосновения молекул тропоколлагена развиваются особые условия преломления, при световой микроскопии наблюдается поперечная исчерченность с периодичностью около 700 Å . Коллагеновые волокна (CF) представляют собой пучки коллагеновых фибрилл, выровненных таким образом, что волокна также демонстрируют поперечные полосы с периодичностью 700Å. В ткани волокна обычно располагаются пучками. По мере созревания коллагеновых волокон между молекулами тропоколлагена образуются ковалентные поперечные связи, что приводит к возрастному снижению растворимости коллагена.
Цементобласты и остеобласты -это клетки, которые также обладают способностью вырабатывать коллаген.
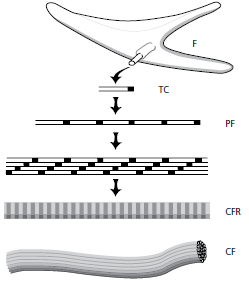
Ретикулярные волокна, как видно на этой микрофотографии (рис. 43), проявляют аргирофильные окрашивающие свойства и довольно многочисленны в ткани, прилегающей к основанию базальной мембраны (стрелки). Однако ретикулярные волокна также встречаются в большом количестве в рыхлой соединительной ткани, окружающей кровеносные сосуды. Таким образом, ретикулярные волокна присутствуют на границе эпителий–соединительная ткань и эндотелий–соединительная ткань.
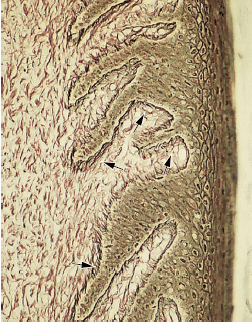
Окситалановые волокна редки в десне, но многочисленны в периодонтальной связке. Они состоят из длинных тонких фибрилл диаметром около 150 Å. Эти соединительнотканные волокна могут быть продемонстрированы при световой микроскопии только после предварительного окисления уксусной кислотой. Микрофотография (рис. 44) иллюстрирует окситалановые волокна (стрелки) в периодонтальной связке, где они имеют ход преимущественно параллельно длинной оси зуба. Функция этих волокон пока неизвестна. Цемент виден слева, а альвеолярная кость-справа.
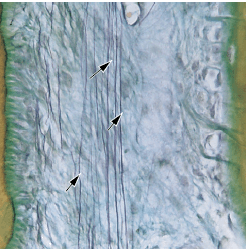
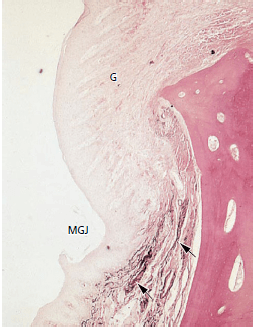
Эластические волокна в соединительной ткани десны и периодонтальной связки присутствуют только в ассоциации с кровеносными сосудами. Однако, как видно на микрофотографии (рис. 45), собственная пластинка и подслизистая оболочка альвеолярной слизистой оболочки содержат многочисленные эластические волокна (стрелки). Десна (G) видимая со стороны мукогингивального соединения (MGJ) не содержит эластических волокон, кроме как в ассоциации с кровеносными сосудами.
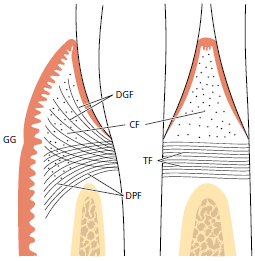
Хотя многие коллагеновые волокна в десне и периодонтальной связке распределены неравномерно или хаотично, большинство из них, как правило, расположены в группах пучков с четкой ориентацией. В соответствии с их расположением и ходом в ткани, ориентированные пучки в десне, можно разделить на следующие группы:
- Круговые волокна (CF) это пучки волокон, которые проходят свой путь в свободной десне и окружают зуб манжеткой или кольцом.
- Зубо-десневые волокна (DGF) внедряются в цемент надальвеолярной части корня и выходят из него веерообразно в свободную десневую ткань вестибулярной, язычной и межпроксимальной поверхностей.
- Зубо-периостальные волокна (DPF) внедряются в ту же часть цемента, что и зубодесневые волокна, но проходят свой путь апикально над вестибулярным и язычным костным гребнем и заканчиваются в ткани прикрепленной десны. В пограничной зоне между свободной и прикрепленной десной, эпителию часто не хватает поддержки со стороны нижележащих ориентированных пучков коллагеновых волокон. В этой области часто присутствует свободная десневая борозда (GG).
- Транссептальные волокна (ТF), видные на рисунке 46, проходят между надальвеолярным цементом соседних зубов. Транссептальные волокна проходят прямо через межзубную перегородку и встраивается в цемент соседних зубов.
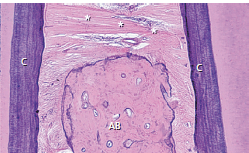
Проиллюстрирована (рис. 47) в гистологическом разрезе ориентация транссептальных волокон (звездочек) в надальвеолярной части межзубной области. Следует отметить также, что помимо соединения цемента (С) соседних зубов, транссептальные волокна также соединяют надальвеолярный цемент (С) с гребнем альвеолярной кости (АВ). Четыре группы пучков коллагеновых волокон укрепляют десну и обеспечивают ее упругость и тонус, необходимые для сохранения архитектуры и целостности зубодесневого крепления.
Межклеточное вещество (матрикс)
Матрикс соединительной ткани вырабатывается главным образом фибробластами, хотя некоторые компоненты вырабатываются тучными клетками, а другими структурами, поступающими из крови.
Матрикс или межклеточное вещество — это среда, в которой находятся клетки соединительной ткани, и она необходима для поддержания нормальной функции соединительной ткани. Таким образом, происходит транспортировка воды, электролитов, питательных веществ, метаболитов и т. д. в отдельные клетки соединительной ткани и из них внутрь матрикса. Основными составляющими соединительнотканного матрикса являются белково–углеводные макромолекулы. Эти комплексы обычно делятся на протеогликаны и гликопротеины. Протеогликаны содержат гликозаминогликаны в виде карбогидратных звеньев (гиалурона сульфат, гепаран сульфат и др.), которые присоединяются к одной или нескольким белковым цепям посредством ковалентных связей. Углеводный компонент всегда преобладает в составе протеогликанов.
Гликозаминогликан, называемый гиалуронан или “гиалуроновая кислота”, вероятно, не связана с белком. Гликопротеины (фибронектин, остеонектин и др.) также содержат полисахариды, но эти макромолекулы отличаются от гликозаминогликанов. Белковый компонент преобладает в составе гликопротеинов. В макромолекулах моно‐ или олигосахариды соединены с одной или несколькими белковыми цепями посредством ковалентных связей.
На рисунке 48 показана данная структура. Нормальная функция соединительной ткани зависит от наличия протеогликанов и гликозаминогликанов. Углеводные фрагменты протеогликанов, гликозаминогликанов, представляют собой большие, гибкие цепочки отрицательно заряженных молекул, каждая из которых занимает довольно большое пространство (а). В такое пространство могут быть включены более мелкие молекулы, например вода и электролиты, в то время как более крупные молекулы не могут войти (b). Таким образом, протеогликаны регулируют диффузию и поток жидкости через матрикс и являются важными детерминантами содержания жидкости в тканях и поддержания осмотического давления. Другими словами, протеогликаны действуют как молекулярный фильтр и, кроме того, играют важную роль в регуляции клеточной миграции (движения) в ткани. Благодаря своей структуре и гидратации макромолекулы сопротивляются деформации, тем самым служат регуляторами консистенции соединительной ткани (c). Если десна под давлением, макромолекулы деформируются.
Когда давление устраняется, макромолекулы восстанавливают свою первоначальную форму. Таким образом, макромолекулы имеют важное значение для упругости десны.
Эпителиально–мезенхимальное взаимодействие
Во время эмбрионального развития различных органов, происходит взаимное индуктивное влияние между эпителием и соединительной тканью. Характерным примером этого явления является развитие зубов. Соединительная ткань является, с одной стороны, определяющим фактором для нормального развития зубной почки, а с другой стороны, эпителий эмали оказывает определенное влияние на развитие мезенхимальных компонентов зубов.
Было высказано предположение, что на дифференцировку тканей во взрослом организме могут влиять факторы окружающей среды. Кожа и слизистые оболочки ,например, часто демонстрируют повышенную кератинизацию и гиперплазию эпителия в областях, которые подвергаются механическому воздействию. Таким образом, ткани, по-видимому, приспосабливаются к внешним раздражителям. Считается, что наличие ороговевшего эпителия на слизистой оболочке жевательного аппарата является адаптацией к механическому раздражению, вызванному жеванием. Однако исследования показали, что характерные особенности эпителия в таких областях определяются генетически. Некоторые соответствующие наблюдения представлены на следующих изображениях
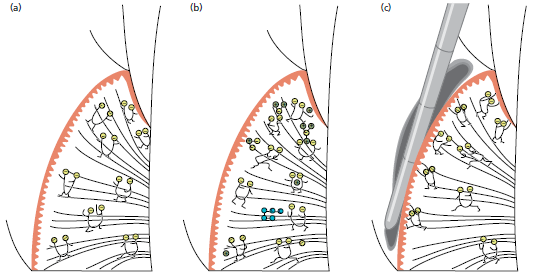
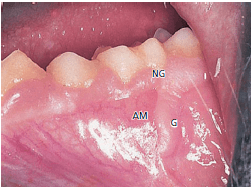
На рис. 49 показан фрагмент трансплантата у обезьяны. Десна (G) и слизистая оболочка альвеол (AM) были перенесены хирургическим путем. Слизистая оболочка альвеол находится в тесном контакте с зубами, в то время как десна расположена в области альвеолярной слизистой оболочки.
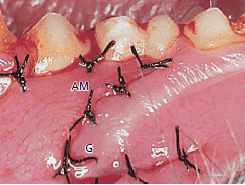
На рис. 50 показана та же область, что и на рис. выше, 4 месяца спустя. Несмотря на то, что пересаженная десна (G) подвижна по отношению к подлежащей кости, как и альвеолярная слизистая оболочка (АМ), она сохранила свои характерные морфологические признаки жевательной слизистой оболочки. Узкая зона новой ороговевшей десны (NG) образовалась между слизистой оболочкой альвеол и зубами.
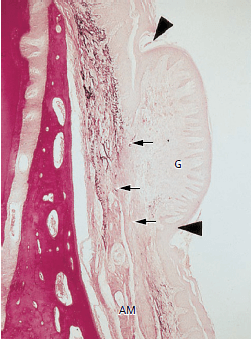
На рис. 51 показан гистологический разрез пересаженной десны, показанный на рис. выше. Так как эластические волокна отсутствуют в соединительной ткани десны, но их много (маленькие стрелки) в соединительной ткани альвеолярной слизистой оболочки (АМ), пересаженная десневая ткань может быть легко идентифицирована. Эпителий, покрывающий пересаженную десневую ткань, имеет на поверхности отчетливый кератиновый слой (между кончиками стрелок), а конфигурация взаимодействия эпителий–соединительная ткань (т. е. ретикулярные колышки и соединительнотканные сосочки) аналогична конфигурации нормальной, непересаженной десне. Таким образом, гетеротопически расположенная десневая ткань сохранила свою первоначальную специфичность. Это наблюдение демонстрирует, что характеристики десны определяются генетически, а не являются результатом функциональной адаптации к окружающей среде.
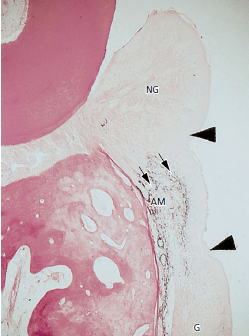
На рис. 52 показан гистологический разрез через коронарную часть области трансплантации, показанный на рис. выше. Пересаженная десневая ткань (G) видна в нижней части микрофотографии. Пересаженная альвеолярная слизистая оболочка (АМ) видна между кончиками стрелок в середине микрофотографии. После операции трансплантат альвеолярной слизистой оболочки располагался в тесном контакте с зубами. После заживления узкая зона ороговевшей десны (NG) развивалась по направлению к альвеолярному трансплантату слизистой оболочки . Эта новая зона десны (NG), которая видна в верхней части гистологического среза, покрыта ороговевшим эпителием, а соединительная ткань не содержит окрашенных фиолетовым эластических волокон. Кроме того, важно отметить, что соединение между ороговевшим и не ороговевшим эпителием (наконечники стрелок) точно соответствует соединению между “эластичной” и “неэластичной” соединительной тканью (маленькая стрелка). Соединительная ткань новой десны восстанавливалась из соединительной ткани надальвеолярной и периодонтальной связок и отделила альвеолярный слизистый трансплантат (AM) от зуба. Вполне вероятно, что эпителий, покрывающий новую десну, мигрировал из соседнего эпителия альвеолярной слизистой оболочки. Это указывает на то, что именно соединительная ткань определяет качество эпителия.
На рис. 53 показан схематический рисунок развития новой узкой зоны ороговевшего десны (NG).
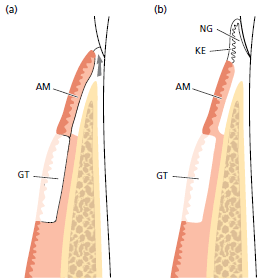
здесь: а — грануляционная ткань (GТ) пролиферировала коронально вдоль поверхности корня (стрелка) и отделила трансплантат альвеолярной слизистой оболочки (АМ) от его первоначального контакта с поверхностью зуба.
b — Эпителиальные клетки мигрировали из альвеолярного трансплантата слизистой оболочки (АМ) во вновь образованную десневую соединительную ткань (NG).
Таким образом, новообразованная десна покрылась ороговевшим эпителием (КЕ), который возник из некератинизированного эпителия альвеолярной слизистой оболочки (АМ). Это означает, что новообразованная соединительная ткань десны обладает способностью индуцировать изменения в дифференцировке эпителия, происходящие из альвеолярной слизистой оболочки. Этот эпителий, который обычно не кератинизирован, по‐видимому, отличается от ороговевшего эпителия из — за стимулов, возникающих из новообразованной соединительной ткани десны. (GТ, десневой трансплантат).
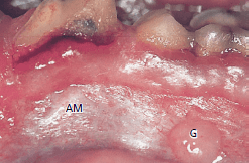
На рис. 54 показана часть десневой соединительной ткани (G) и альвеолярной соединительной ткани слизистой оболочки (AM), которая после трансплантации срослась в раневыми участками на альвеолярной слизистой оболочке. Эпителизация этих трансплантатов может происходить только за счет миграции эпителиальных клеток из окружающей альвеолярной слизистой оболочки.
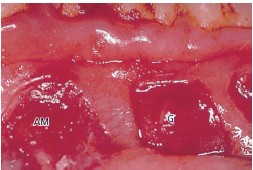
На рис. 55 показана пересаженная десневая соединительная ткань (G) после повторной эпителизации. Эта часть ткани приобрела вид, похожий на вид нормальной десны, что указывает на то, что эта соединительная ткань теперь покрыта ороговевшим эпителием. Пересаженная соединительная ткань из альвеолярной слизистой оболочки (АМ) покрыта некератинизированным эпителием и имеет тот же внешний вид, что и окружающая альвеолярная слизистая оболочка.
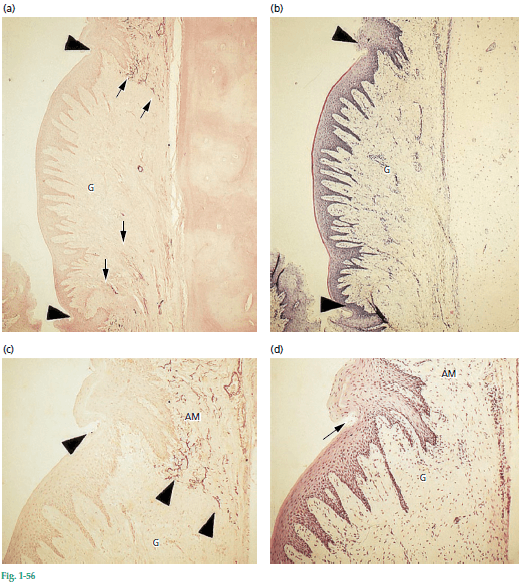
На рис. 56 показаны два гистологических среза через область пересаженной десневой соединительной ткани. Участок, показанный на а, окрашен за счет эластичных волокон (стрелки). Ткань в середине без эластических волокон-это пересаженная десневая соединительная ткань (G). На рис. b показан соседний участок, окрашенный гематоксилином и эозином. При сравнении рис. а и b, видно, что пересаженная десневая соединительная ткань покрыта ороговевшим эпителием (между наконечниками стрелок), а также что граница эпителий–соединительная ткань имеет тот же волнистый ход (т. е. ретикулярные колышки и соединительнотканные сосочки), что и в нормальной десне.
Микрофотографии показанные на рис. с и рис. d иллюстрирует, при более высоком увеличении, пограничную область между альвеолярной слизистой оболочкой (AM) и пересаженной десневой соединительной тканью (G). Обратите внимание на отчетливую взаимосвязь между ороговевшим эпителием (стрелка) и “неэластичной” соединительной тканью (наконечники стрел), а также между некератинизированным эпителием и “эластичной” соединительной тканью. Установление таких тесных взаимосвязей во время заживления подразумевает, что пересаженная десневая соединительная ткань обладает способностью изменять дифференцировку эпителиальных клеток, как предполагалось ранее. Клетки эпителия альвеолярной слизистой оболочки, начав как некератинизующиеся клетки, очевидно, стали кератинизирующими клетками. Это означает, что специфичность эпителия десны определяется генетическими факторами, присущими соединительной ткани.
Периодонтальная связка
Периодонтальная связка — это мягкая, богатая сосудами и клетками соединительная ткань, которая окружает корни зубов и соединяет корневой цемент со стенкой альвеолы.
В направлении коронки периодонтальная связка непрерывно соединена с собственной пластинкой десны и отделена от десны пучками коллагеновых волокон, которые соединяют гребень альвеолярной кости с корнем (волокна альвеолярного гребня).
На рис. 57 представлена рентгенограмма премолярно–молярной области нижней челюсти.
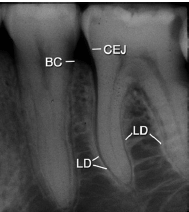
На рентгенограммах можно выделить два типа альвеолярной кости:
- Часть альвеолярного отростка, которая покрывает альвеолы, обозначаемая как “пластинка твердой мозговой оболочки” (LD).
- Участок альвеолярного отростка, который на рентгенограмме имеет вид сетчатой структуры, обозначается как “трабекулярная кость”.
Периодонтальная связка расположена в пространстве между корнями зубов и твердой пластинкой (LD) или собственно альвеолярная кость. Альвеолярная кость окружает зуб от вершины до уровня приблизительно 1 мм апикальнее цементоэмалевого соединения (CEJ). Корональная граница кости называется костным гребнем (BC).
Периодонтальное связочное пространство имеет форму песочных часов и самые узкая часть на уровне среднего корня. Ширина периодонтальной связки составляет приблизительно 0,25 мм (диапазон 0,2–0,4 мм). Наличие периодонтальной связки позволяет силам, возникающим во время жевательной функции и других контактов зубов, распределяться альвеолярным отростком через собственно альвеолярную кость. Периодонтальная связка также необходима для подвижности зубов.
Подвижность зубов в значительной степени определяется шириной, высотой и качеством периодонтальной связки.
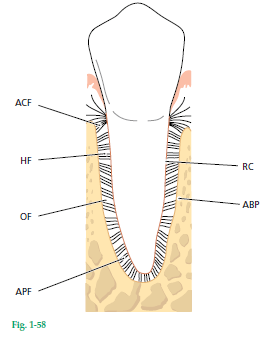
На рис. 58 схематично показано, как периодонтальная связка располагается между собственно альвеолярной костью (ABP) и корневым цементом (RC). Зуб соединен с костью пучками коллагеновых волокон, которые по своему расположению можно разделить на следующие основные группы:
- Волокна альвеолярного гребня (АCF)
- Горизонтальные волокна (HF)
- Косые волокна (OF)
- Апикальные волокна (АPF).
Периодонтальная связка и корневой цемент развиваются из рыхлой соединительной ткани (фолликула), которая окружает почку зуба. На схематическом рисунке показаны различные стадии в организации периодонтальной связки, которая формируется одновременно с развитием корня и прорезыванием зуба.
Рисунок 59:
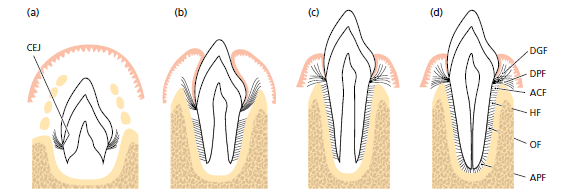
здесь: а — зубная почка формируется в крипте кости. Коллагеновые волокна, продуцируемые фибробластами в рыхлой соединительной ткани вокруг бутона зуба, внедряются в процессе их созревания в составе вновь сформированного цемента сразу апикальнее цементо-эмалевого соединения (CEJ). Эти пучки волокон, ориентированные к коронарной части костной крипты, впоследствии образуют группу зубодесневых волокон, и группу транссептальных волокон, которые относятся к ориентированным волокнам десны.
b — истинные волокна периодонтальной связки, главные волокна, развиваются при прорезывани зуба. В начале эти волокна могут быть идентифицированы при их входе в самую краевую часть альвеолярной кости.
с — видны более апикально расположенные пучки ориентированных коллагеновых волокон.
d — ориентация пучков коллагеновых волокон непрерывно изменяется в течение фазы прорезывания зуба. Во‐первых, когда зуб достиг контакта в окклюзии и функционирует нормально, волокна периодонтальной связки объединяются в группы хорошо ориентированных зубочелюстных коллагеновых волокон. Эти коллагеновые структуры подвергаются постоянному ремоделированию (т. е. рассасыванию старых волокон и образованию новых). (DGF зубодесневые волокна; DPF зубопериостальных волокна; HF горизонтальные волокона; OF косые волокона; АPF верхушечнык волокона).
Этот схематический рисунок 60 иллюстрирует развитие основных волокон периодонтальной связки. Собственно альвеолярная кость (ABP) видна слева, периодонтальная связка (PL) — в центре, а корневой цемент (RC) — справа.
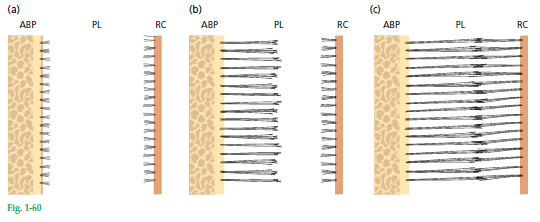
а — сначала обнаруживаются мелкие, тонкие, щеткообразные фибриллы, возникающие из корневого цемента и выступающие в периодонтальное пространство. На этой стадии поверхность кости покрывается остеобластами. С поверхности кости видно лишь небольшое их количество излучающее тонкие коллагеновые фибриллы.
b — далее количество и толщина волокон, входящих в кость, увеличиваются. Эти волокна расходятся в сторону рыхлой соединительной ткани в средней части пародонтальной связки, которая содержит более или менее хаотично ориентированные коллагеновые фибриллы. Волокна, выходящие из цемента, все еще короткие, а те, что входят в кость, постепенно удлиняются. Концевые части этих волокон имеют пальцевидные выступы.
с — волокна, происходящие из цемента, впоследствии увеличиваются в длину и толщину и сливаются в пародонтальном пространстве связок с волокнами, происходящими из альвеолярной кости. Когда зуб после прорезывания достигает контакта в окклюзии и начинает функционировать, основные волокна организуются в пучки и непрерывно проходят от кости к цементу.
Рисунок 60:

здесь: а — показано, как основные волокна периодонтальной связки (PDL) непрерывно проходят от корневого цемента до собственно альвеолярной кости (ABP). Основные волокна, встроенные в цемент (Шарпеевские волокна), имеют меньший диаметр, но более многочисленны, чем те, которые встроены в собственно альвеолярную кость.
b — показана поляризованная версия а. На этом изображении видны Шарпеевские волокна (SF) , пронизывающие не только цемент (C), но и всю ширину собственно альвеолярной кости (ABP). Периодонтальная связка также содержит несколько эластичных волокон, связанных с кровеносными сосудами. Окситалановые волокна также присутствуют в периодонте. Они имеют преимущественно апико‐окклюзионную ориентацию и располагаются в связке ближе к зубу, чем к альвеолярной кости. Очень часто их вставляют в цемент. Их функция еще не определена.
Клетками периодонтальной связки являются: фибробласты, остеобласты, цементобласты, остеокласты, а также эпителиальные клетки и нервные волокна. Фибробласты распределены вдоль основных волокон, в то время как цементобласты по поверхности цемента, а остеобласты по поверхности кости.
Рис. 62. а — показано наличие скоплений эпителиальных клеток (ER) в периодонтальной связке (PDL). Эти клетки, называемые остатками эпителиальных клеток Маллассе, представляют собой остатки эпителиальной корневой оболочки Хертвига. Остатки эпителиальных клеток располагаются в периодонтальной связке на расстоянии 15–75 мкм от цемента (С) на поверхности корня. Группа таких эпителиальных клеток хорошо видна на примере рис.b.
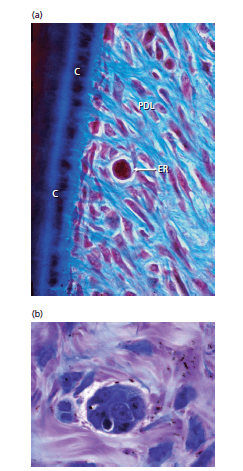
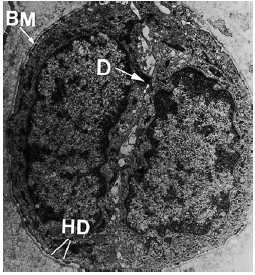
Под электронным микроскопом (рис. 63) видно, что остатки эпителиальных клеток окружены базальной мембраной (BM) и что мембраны эпителиальных клеток имеют десмосомы (D), а также гемидесмосомы (HD). Эпителиальные клетки содержат лишь несколько митохондрий и имеют слабо развитый эндоплазматический ретикулум. Это означает, что они являются живыми, но неработающими клетками с незначительным метаболизмом.
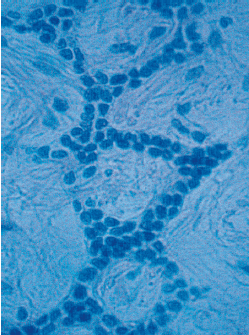
Представлена микрофотография (рис. 64) периодонтальной связки удаленного зуба. Этот образец показывает, что остатки эпителиальных клеток Малассе, которые в обычных гистологических срезах появляются в виде изолированных групп, по сути, образуют сплошную сеть эпителиальных клеток, окружающих корень. В настоящее время их назначение неизвестно.
Корневой цемент
Цемент-это специализированная минерализованная ткань, покрывающая поверхность корней и, иногда, небольшие участки коронки зубов. Он также может распространяться в корневой канал. В отличие от кости, цемент не содержит кровеносных или лимфатических сосудов, не имеет иннервации, не подвергается физиологической резорбции или ремоделированию, но характеризуется непрерывным отложением в течение всей жизни. Как и другие минерализованные ткани, он содержит коллагеновые фибриллы, встроенные в органическую матрицу. Его минеральный состав представлен в основном, гидроксиапатитом (около 65% по весу, немного больше, чем у кости -60%). Цемент выполняет различные функции. Он прикрепляет основные волокна периодонтальной связки к корню и способствует процессу восстановления после повреждения поверхности корня. Он также играет важную роль при изменеии положения зуба.
Различают формы цемента:
- Бесклеточный афибриллярный цемент (ААС) находится в основном в пришеечной части эмали.
- Бесклеточный наружный волокнистый цемент (AEFC) находится в корональной и средней частях корня и содержит в основном пучки волокон Шарпея. Этот тип цемента является важной частью аппарата прикрепления, соединяя зуб с костью (собственно альвеолярной костью).
- Клеточный смешанный слоистой цемент (CMSC) располагается в верхушечной трети корней и в зонах фуркации. Он содержит как внешние, так и внутренние волокна, а также цементоциты.
- Клеточный внутренний волокнистый цемент (CIFC) находится главным образом в резорбционных лакунах и содержит внутренние волокна и цементоциты.
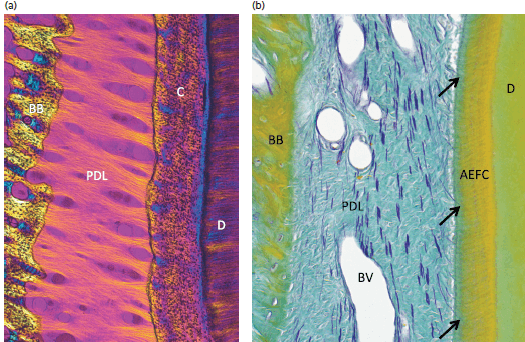
На рис. 65: а — показан препарат в поляризованном свете. Основные коллагеновые волокна периодонтальной связки (PDL) проходят между корнем, покрытым цементом (C), и альвеолярным отростком, покрытым пучками костной ткани (BB). Части основных волокон периодонтальной связки, которые встроены в корневой цемент и в кость называются Шарпеевскими волокнами (D, дентин).
b — окситалановые волокна в периодонтальной связке (PDL) проходят в апикально-корональном направлении; некоторые (стрелки) входят в бесклеточный наружный волокнистый цемент (AEFC). Многие окситалановые волокна видны вокруг кровеносных сосудов (BV) в периодонтальной связке. Окситалановые волокна могут выполнять функцию передачи механического давления между корнем зуба и периодонтальной связкой. (ВВ, костная стенка лунки; D, дентин).
На рис. 66:
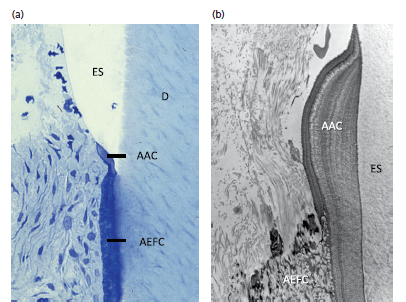
а — показано присутствие бесклеточного афибриллярного цемента (ААС) в области дентино-цементного соединения. Бесклеточный афибриллярный цемент покрывает незначительные участки пришеечной эмали. Он не содержит ни клеток, ни коллагеновых фибрилл. Он может образовывать изолированные участки на эмали или быть соединенным с бесклеточным наружный волокнистым цементом (AEFC). Бесклеточный афибриллярный цемент может образовываться, когда редуцированный эпителий эмали очагово дезинтегрируется так, что открытая поверхность эмали вступает в контакт с окружающей мягкой соединительной тканью (D, дентин; ES, эмалевое пространство).
b — показана морфология бесклеточного афибриллярного цемента (ААС) под электронным микроскопом. Бесклеточный афибриллярный цемент простирается от бесклеточного наружного волокнистого цемента (AEFC) в корональном направлении. Слоистый вид бесклеточного афибриллярного цемента указывает на периоды активного отложения и покоя. Функция бесклеточного афибриллярного цемента неясна. Умеренно электронно-плотный материал в эмалевом пространстве (ES), прилегающий к бесклеточному афибриллярному цементу представляет собой постоянный эмалевый матрикс.
Показаны (рис. 67) три стадии развития бесклеточного наружного волокнистого цемента (AEFC). Бесклеточный наружный волокнистый цемент образуется одновременно с образованием корневого дентина. В начале развития корня эпителиальная оболочка Хертвига, которая выстилает новообразованный предентин, фрагментируется. Затем цементобласты начинают синтезировать коллагеновые волокна большого размера, которые имплантируются под прямым углом к поверхности. Во время непрерывного формирования бесклеточного наружного волокнистого цемента, части коротких коллагеновых волокон, прилегающих к корню, внедряются в минерализованную ткань.
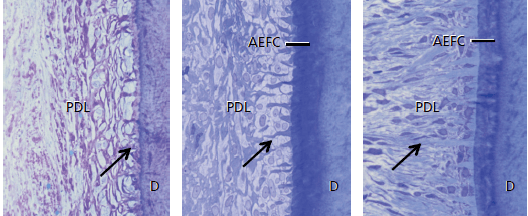
здесь: а — показаны короткие коллагеновые волокна (стрелка), выступающие с поверхности дентина (D) в периодонтальную связку (PDL), которые составляют будущие волокна Шарпея. Однако слой цемента еще не виден.
b — показаны короткие коллагеновые волокна (стрелка), выступающие с поверхности корней, но их основания теперь встроены, как волокна Шарпея, в минерализованный цемент.
с — показано, что большинство коллагеновых волокон удлиняются и продолжаются в пародонтальном пространстве связки. Эти микрофотографии показывают, что волокна Шарпея в цементе являются прямым продолжением основных волокон периодонтальной связки и надальвеолярной соединительной ткани. AEFC увеличивается в течение всей жизни с очень медленной скоростью роста 1,5–4,0 мкм/год.
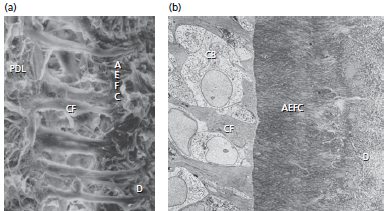
На рис. 68: а — показана электронная микрофотография недекальцинированной поверхности бесклеточного наружного волокнистого цемента (АEFC). Заметим, что внешние волокна прикрепляются к дентину (D), пересекают минерализованный слой цемента в виде волокон Шарпея и соединяются с коллагеновыми волокнами (CF) периодонтальной связки (PDL).
b — показана электронная микрофотография бесклеточного наружного волокнистого цемента (AEFC). Волокна Шарпея (т. е. внешние коллагеновые волокна бесклеточного наружного волокнистого цемента) проходят от поверхности дентина (D) через минерализованный цементный слой и продолжают выходить за пределы цемента в качествеосновных коллагеновых волокон (CF) в периодонтальную связку. Цементобласты (CB) занимают промежутки между выступающими коллагеновыми волокнами.
На рис. 69: a — показана электронная микрофотография бесклеточного наружного волокнистого цемента (АEFC) в момоент минерализации. Волокна Шарпея покидают цемент и продолжаются как основные волокна периодонтальной связки. Цементобласты (CB) занимают пространство между плотно упакованными коллагеновыми фибриллами. Характерная поперечная исчерченность коллагеновых фибрилл маскируется в цементе из — за присутствия неколлагеновых белков. Минерализация происходит путем осаждения кристаллов гидроксиапатита сначала в коллагеновых волокнах, затем на поверхности волокон и, наконец, в межфибриллярной матрице.
b — показан бесклеточный наружный волокнистого цемента (AEFC) в момент минерализации. Срез ткани обрабатывали антителами к костному сиалопротеину. Этот неколлагеновый протеин выполняет функцию регуляции минерализации твердых тканей на основе коллагена. Частицы золота маркируют межфибриллярный матрикс минерализованного цемента, в то время как немаскированные коллагеновые фибриллы, которые покидают цемент и распространяются в пародонтальное пространство связок, не маркируются.

На рис. 70:
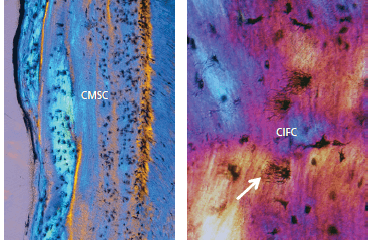
а — показан неокрашенный и недекальцинированный участок ткани в поляризованном свете. На микрофотографии показана структура клеточного смешанного слоистого цемента (CМSC), состоящего из чередующихся слоев бесклеточного наружного волокнистого цемента и клеточного внутреннего волокнистого цемента. В отличие от бесклеточного наружного волокнистого цемента, клеточный внутренний волокнистый цемент содержит клетки и внутренние волокна. В то время как внешние волокна Шарпея пересекают слой цемента и покидают его при минерализации, внутренние волокна полностью находятся внутри цемента. Клетки, входящие в состав цемента, называются цементоцитами. Клеточный смешанный слоистый цемент закладывается на протяжении всего функционального периода зуба. Расслоение клеточного смешанного слоистогоо цемента обычно нерегулярно. Он встречается на срединной и апикальной поверхностях корней, а также в зонах фуркации. Цемент становится значительно шире в апикальной части корня, по сравнению с пришеечной областью. В апикальной части корня цемент часто имеет ширину 150-250 мкм или даже больше. Он часто содержит инкрементные линии, указывающие на чередующиеся периоды формирования и покоя.
b — показан неокрашенный и недекальцинированный участок ткани в поляризованном свете. Цементоциты (черные клетки) находятся в лакунах в клеточном внутреннем волокнистом цементе (CIFC), который находится в клеточном смешанном слоистом цементе. Цементоциты взаимодействуют друг с другом через сеть цитоплазматических отростков (стрелка), проходящих через канальцы в цементе. Большинство клеточных отростков располагаются на поверхности цемента (слева). Цементоциты также связываются с цементобластами на поверхности посредством цитоплазматическихотростков. Наличие цементоцитов позволяет транспортировать питательные вещества и продукты жизнедеятельности через цемент и способствует поддержанию жизнедеятельности этой минерализованной ткани.
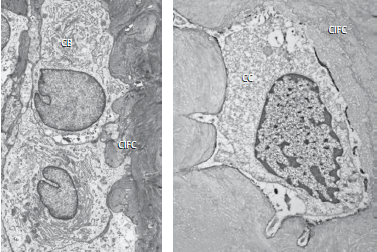
На рис. 71: а — показана электронная микрофотография с поверхности клеточного внутреннего волокнистого цемента (CIFC). Цементоид выстлан типичными цементобластами (CB). Это крупные кубовидные клетки с круглым, богатым эухроматином ядром. Изобилие грубого эндоплазматического ретикулума указывает на то, что эти клетки очень активны и продуцируют белки, которые секретируются во внеклеточное пространство. Они формируют цементоидный пласт, состоящий из коллагеновой матрицы, которая позже минерализуется. Как правило, бесклеточный наружный волокнистый цемент более минерализован, чем клеточный смешанный слоистый цемент и клеточный внутренний волокнистый цемент. Иногда минерализуется только периферия волокон Шарпея клеточного смешанного слоистого цемента, оставляя внутри волокон неминерализованное ядро.
b — представлена электронная микрофотография, которая показывает цементоцит (CC) в клеточном внутреннем волокнистом цементе (CIFC). Цементоциты- это цементобласты, которые попали в ловушку цементного матрикса. Они располагаются в лакунах, из которых идут несколько канальцев, пересекая цементный матрикс и сообщаясь с соседними цементоцитами. Лакуны цементоцитов в более глубоких частях цемента часто оказываются пустыми, что может быть связано с превышением критического расстояния для обмена веществ.
Продолжение статьи «Строение пародонта: анатомия и гистология тканей пародонта. Часть 3».
Перевод материала «Anatomy of Periodontal Tissues» Jan Lindhe, Thorkild Karring, Maurício Araújo выполнен специально для проекта «Современная пародонтология».